
Przegląd Ewassa Oldupy w wąwozie Olduvai (Oldupai) w Tanzanii. Michał Petraglia. [ https://www.archaeology.wiki/blog/2021/01/08/hominins-of-olduvai-gorge-coped-with-ecological-changes/ ]
Streszczenie
Szybkie zmiany środowiskowe są katalizatorem ewolucji człowieka, napędzającym innowacje dietetyczne, dywersyfikację siedlisk i rozproszenie. Jednak brakuje informacji, aby ocenić adaptacje homininów do zmieniającej się fizjografii podczas kluczowych etapów ewolucji, takich jak wczesny plejstocen. Tutaj przedstawiamy zbiór danych multiproxy z Ewass Oldupa, w zachodnim plio-plejstoceńskim zagłębieniu ryftowym wąwozu Olduvai (obecnie Oldupai), Tanzania, aby rozwiązać tę lukę i zaoferować ekologiczną perspektywę na temat adaptacji człowieka dwa miliony lat temu. Najwcześniejsze homininy Oldupai zamieszkiwały kolejno równiny zalewowe krętych kanałów, a następnie konteksty pod wpływem rzeki, które obecnie obejmują najstarsze udokumentowane regionalnie środowisko paleojeziorne. Wczesne narzędzia oldowańskie ujawniają jednorodną technologię wykorzystywania zróżnicowanych, szybko zmieniających się środowisk, od łąk paprociowych po mozaiki leśne, naturalnie wypalone krajobrazy, po nadbrzeżne lasy/gaje palmowe, a także hiperkseryczne stepy. Homininy okresowo wykorzystywały pojawiające się krajobrazy i biomy zaburzeń wielokrotnie w ciągu 235 000 lat, wyprzedzając tym samym o ponad 180 000 lat najwcześniejsze znane homininy i przemysły oldowańskie ze wschodniej strony basenu.
Wstęp
Wczesnym plejstocenie u homininów zaszły duże przemiany biologiczne, którym towarzyszyło zwiększone uzależnienie od narzędzi kamiennych1 , ogólna dywersyfikacja diety2 , 3 , 4 , 5 , 6 i dalekosiężne rozprzestrzenianie7 , 8 , 9. Kontekst behawioralny tych zmian pozostaje trudny do uchwycenia ze względu na brak zestawów danych chronostratygraficznych i środowiskowych o wysokiej rozdzielczości w bezpośrednim powiązaniu ze szczątkami kultury oldowajskiej, dostępnych jedynie dla niewielkiej liczby stanowisk10 , 11 , 12 , 13 , 14 , 15 , 16 , 17 . Ponadto ekstrapolacja zewnętrznych danych paleoekologicznych z odwiertów wykonanych w tym samym czasie co jezioro oraz sekwencji wierceń w jeziorach18 ma ograniczoną przydatność w zrozumieniu lokalnego użytkowania gruntów, synchronicznego/diachronicznego zajmowania zróżnicowanych środowisk lądowych i docelowej eksploatacji siedlisk przez człowiekowatych oldowańskich.
Rekonstrukcja paleośrodowiskowa ze stanowisk 2,6 do 1,9 Ma opierała się na pośrednich przybliżeniach dawnej roślinności na podstawie fauny i/lub stabilnych izotopów 13 , 14 , 15 , 16 , 17 , 19 , 20 , oznaczających zmiennie otwarte mozaiki trawiaste i leśne w środowiskach rzecznych, często z ograniczonych interwałów stratygraficznych 21 , 22 . Nasze badania godzą najwcześniejsze narzędzia kamienne olduwajskie z wąwozu Oldupai, kluczowego kompleksu do badania sposobów życia homininów 23 , 24 , z wieloproksymacyjnymi zestawami danych w bezpośrednim powiązaniu ze stratyfikowanymi zespołami archeologicznymi i kopalnymi, dokumentującymi epizodyczną eksploatację tego samego miejsca w zróżnicowanych kontekstach geomorfologicznych i osadach sedymentacyjnych. Ten zestaw danych stanowi model badań interdyscyplinarnych mających na celu wyjaśnienie kontekstu środowiskowego wczesnych stanowisk olduwajskich. W niniejszym opracowaniu badamy zachowanie człowiekowatych w powiązaniu ze społecznościami fauny i roślin oraz przedstawiamy dowody na fizjonomię roślinności i jej pokrycie na podstawie analizy fitolitów i palinologii, wartości izotopowych n -alkanów z wosków roślinnych, stabilnych izotopów ze szkliwa oraz regionalnych pożarów na podstawie stężeń mikrowęgla drzewnego.
Wyniki
Stratygrafia i archeologia
Stanowisko Ewass Oldupa (lokalizacja geologiczna 63, ryc. 1 ) 23 znajduje się 350 m na północny wschód od lokalizacji geologicznej 64, gdzie zachowano kompletną szczękę górną i dolną powierzchnię Homo habilis (OH 65) 22 z warstw datowanych na ~1,82 Ma 25 , 26 . W Ewass Oldupa odsłoniliśmy grubą sekwencję osadową z wiekiem ograniczonym istniejącymi datami 40 Ar/ 39 Ar tufów odciskanych geochemicznie 26 , 27, którą dodatkowo ograniczono do sześciu lokalizacji wzdłuż 2-kilometrowego transektu (rys. uzupełniający 1 , tabela 1 ). Najniższą jednostką stratygraficzną jest ignimbryt Naabi z formacji Ngorongoro 18 (ryc. 1 i 2a ). Powyżej znajduje się wysoce heterogeniczna strefa składu grubego tufu krystalicznego skalenia (CFCTcz) 28 (rys. 1 i 2c ), w tym gruby tuf krystaliczny skalenia (2,015 ± 0,006 Ma) 25 . Dolna warstwa I zaczyna się od osadzania zielonej woskowej gliny 29 i przekładkowych warstw węglanowych, co wskazuje na ekspansję jeziora 1 km dalej na zachód i wystąpienie wcześniej niż wcześniej odnotowano 18 , 26 , 27 , 28 , 29 (rys. 1 – 2e ). Tuf IA (~2,0 Ma) 18 (rys. 1 i 2f, g ) zawiera charakterystyczny bogaty w Mg augit, przerobione ziarna detrytyczne pochodzące z podłoża i materiały CFCTcz 25 , 26 , 27 , 28 . Górne Warstwa I obejmuje Tuf IB (1,848 ± 0,003 Ma) 25 . Bezpośrednio nad nim znajdują się woskowe, zielonobrązowe iłowce osadzone w okresie wysokiego poziomu jeziora (rys. 1 , 2h ). Nad tymi glinami leży słabo uwarstwiony mułowiec o geochemicznym podpisie Tufu IC (1,832 ± 0,003 / 1,848 ± 0,008 Ma) 22 , 25 , 26 . Zamykający go słabo płaski uwarstwiony piaskowiec jest geochemicznie zgodny z Tufem ID, zidentyfikowanym gdzie indziej w zachodnim wąwozie jako przedział zawierający szczątki Homo habilis (OH65) 22 , 26 . Petrograficznie i geochemicznie zidentyfikowano cienką warstwę z grudkami węglanowymi i gruboziarnistym materiałem krzemionkowym jako tuf Ng’eju (1,818 ± 0,006 mln lat temu) 25 , 26.

a Mapa lokalizacji. Symbol gwiazdki oznacza stanowisko Ewass Oldupa. Ostatnio opublikowane otwory wiertnicze 18 są pokazane w celach informacyjnych. Oznaczono również główne typy skał potencjalnie dostępnych do eksploatacji przez homininy w całym regionie. b Pojedynczy przekrój stratygraficzny z Ewass Oldupa, podzielony na dwa segmenty: dolny (i) i górny (ii). Formacja Ngorongoro: sukcesja zaczyna się od Naabi, zielono-szarego, kwarcowo-trachitowego spawanego tufu datowanego na 2,038 ± 0,005 Ma 25 . Cienka czerwona diamiktyt pokrywa Naabi i oddziela go od serii jednostek zdominowanych przez piaskowiec klarujący się ku górze, składających się z maficznego tephry (rys. 2b ), który jest przykryty mułowcem. Formacja Ngorongoro, strefa składowa CFCT: warstwy składają się z czerwonobrązowego piaskowca, detrytusu wulkanicznego i tefry/krzemioklastycznego diamktytu 28 . (i) Dolne złoże I — sześć jednostek, które ku górze drobnoziarniste od konglomeratu otoczakowo-kamienistego do krzyżowo warstwowanego piaskowca przypisuje się meandrującym korytom rzecznym, poniżej tufu IA. Następnie drobnoziarnisty mulisty piaskowiec, który drobnoziarnisty w woskowaty, zielonobrązowy iłowiec z węglanowymi grudkami został osadzony w otoczeniu jeziora. Na koniec przewarstwione warstwy piaskowca, mułowca, iłowca i tufów rejestrują zmienne środowiska od jeziornego do płytkiego brzegu jeziora do równiny zalewowej z małymi kanałami rzecznymi. (ii) Górne złoże I — zaczyna się od tufu IB, leżącego nad woskowatym, zielonobrązowym iłowcem (rys. 2H ), a następnie tufu IC. Uwieńczeniem tej sekwencji jest beżowy, słabo uwarstwiony piaskowiec, który jest geochemicznie zgodny z tufem 1D 22 , 25 , 26 . Powyżej, tuf Ng’eju jest spotykany w sekwencji drobnoziarnistych osadów zalewowych. Dodatkowe informacje stratygraficzne przedstawiono na rys. 5 .
Obraz w pełnym rozmiarze
Ryc. 2: Kluczowe poziomy stratygraficzne.

a Formacja Ngorongoro: ignimbryt Naabi pokrywający podłoże metamorficzne wzdłuż rzeki Oldupai. b Formacja Ngorongoro: piaski mafijne bezpośrednio nad Naabi w wykopie 7. c Formacja Ngorongoro: strefa składowa CFCT, wykop 5, charakteryzująca się licznymi ryzolitami. d Dolne złoże I: zlepieńcowate złoże wypełniające kanał z warstwowaniem poprzecznym koryta nachylonym ku wschodowi. e Dolne złoże I: woskowe iłowce z wykopu 3 przykryte warstwami węglanowymi. f Dolne złoże I: wielopiętrowe osady pasa kanału rzecznego w wykopie 2, kilka metrów pod tufem IA. (Dźwig metryczny na ziemi dla określenia skali.) g Dolne złoże I: odsłonięcie Ewass Oldupa tufu IA i leżących pod nim cienkich warstw tufowych. h Górne złoże I: gruba jednostka woskowego iłowca przykrywająca tuf IB i leżący pod nim tuf IC. Więcej informacji stratygraficznych można znaleźć na rys. 5 .
Obraz w pełnym rozmiarze
Zapis geoarcheologiczny sekwencji Ewass Oldupa pokazuje dowody na obecność człowieka 2,0–1,9 Ma poprzez szybko zmieniające się środowiska depozycyjne, które obejmowały meandrujące strumienie przecinające stożek wulkanoklastyczny, po których następowały zdarzenia przepływu masowego, kilka cykli jeziornych transgresji/regresji, duże i małe postępujące systemy rzeczne z interkalowanym wulkanizmem. Co ważne, ten zapis siedlisk homininów poprzedza, według naszej wiedzy, najstarsze złoża zawierające skamieliny i narzędzia z lokalizacji wschodniego basenu „Douglas Korongo” (DK), które tradycyjnie reprezentowały najwcześniejszą obecność oldowanów w regionie na ~1,848 ± 0,003 Ma 24 , 30 , 31 .
Wykopaliska w Ewass Oldupa ujawniły 1373 okazy skamieniałości i 565 artefaktów kamiennych (Tabela uzupełniająca 1 ), wykazujące spójne cechy technologiczne od okresu bezpośrednio po Naabi do podstawy złoża II. Obecne są wszystkie etapy kruszenia (Tabela uzupełniająca 1 ). Skały kamienne pochodzą z tablicowych, średnioziarnistych, szarawych/białawych płyt kwarcytowych, które są skupione (7/m2 ) w odrębnych zespołach archeostratygraficznych (7/40 m2 ) (rys. 3 i 4 ). Płatki są dominującym produktem (>60%), przy czym sferoidy i materiały perkusyjne są rzadkie. Nie ma zróżnicowanego zarządzania kruszonymi powierzchniami, zatem systemy technologiczne nie podążają za hierarchicznymi strategiami redukcji. Jednokierunkowe i wielokierunkowe kruszenie rdzeni utrzymywano w ramach ograniczeń naturalnych kształtów i kątów uderzeń. Spośród siedmiu metod eksploatacji zarejestrowanych w 58 rdzeniach dominują metody multipolarno-multifacialne (liczba rdzeni = 19, mediana liczby ekstrakcji = 9), unipolarno-podłużne ( n = 15, mediana = 4) i ortogonalnie-dwufacialne ( n = 13, mediana = 5) (rys. uzupełniający 2c ). Intensywność redukcji kamienia jest wnioskowana z minimalnej liczby ekstrakcji na rdzeń, która waha się od dwóch do 16 usunięć. Ogólnie rzecz biorąc, prostota technicznego repertuaru Ewassa Oldupy jest wspólna z innymi wczesnymi zespołami oldowańskimi 6 , 10 , 11 , 12 , 14 , 15 , w tym takie cechy, jak swobodna, twarda perkusja młotka i redukcja multifacialna. Analiza głównych składowych (PCA) (rys. uzupełniający 2a, b , rys. uzupełniający 3 ) pokazuje, że Oldowan Ewassa Oldupy dzieli wiele cech technicznych z Kanjera South 14 , Fejej 11 i do pewnego stopnia z Frida Leakey Korongo Zinj (FLK Zinj) 24 . Co istotne, kilka aspektów przemysłu kamieniarskiego Ewassa Oldupy (rdzenie unifacjalne i bipolarne, liczba wydobyć, rozmiar odłupków, fragmenty kanciaste i niewielka liczba narzędzi udarowych) najlepiej pasuje do profilu techniczno-typologicznego widocznego w starszych stanowiskach Oldowan (Gona 10 i Gona 12) 15 . Ponadto PCA wydobywa mozaikę wczesnych i klasycznych cech oldowańskich w skałach liticznych Ewassa Oldupy, które mieszczą się pomiędzy najstarszymi skałami oldowańskimi > 2 mln lat temu i młodszymi zespołami ≤1,85 mln lat temu, gdzie występuje wiele sferoidów. Podobnie PCA, obejmująca 18 zespołów i obejmująca 11 zmiennych technicznych15 , pokazuje odstający charakter Lomekwi i jego całkowity brak powinowactwa z Ewass Oldupa (rysunki uzupełniające 2 , 3, Tabela uzupełniająca 2 ).
Rys. 3: Mapowanie wykopanych materiałów.

a Wykres wykopanych kamieni i okazów kopalnych. b – d Po prawej: pionowe projekcje artefaktów kamiennych i kości pokazują oddzielne horyzonty archeologiczne (jednostki osi są w metrach). Po lewej: analiza gęstości jądra pokazuje przestrzenne zmiany w akumulacji materiałów archeologicznych (najgęstsze w najciemniejszym błękicie). Pozycja stratygraficzna wykopów 2, 3 i 5 w obrębie całej kolumny jest przedstawiona na ryc. 5 .
Obraz w pełnym rozmiarze
Ryc. 4: Wybór narzędzi kamiennych z Ewass Oldupa.

Formacja Ngorongoro: najwcześniejszy Oldowan, po Naabi, Wykop 7: a Rdzeń kwarcytowy wielobiegunowy-wielopostaciowy. b – c Płatki kwarcytu. Formacja Ngorongoro, strefa składowa Contact CFCT/Dno złoża I, Wykop 5: d Narzędzie do rozdrabniania ignimbrytu. e Rozdrabniacz ignimbrytu. f Rdzeń podłużny unipolarny kwarcytu. g Rdzeń kwarcytowy wielobiegunowy-wielopostaciowy. Dolne złoże I: najwcześniejsza ekspansja jeziora, Wykop 3: h Sferoida kwarcytu. i Płatki kwarcytu. Dolne złoże I: postępujący system rzeczny poniżej tufu IA, Wykop 2: j – l Płatki kwarcytu.
Obraz w pełnym rozmiarze
Porównanie geochemicznego odcisku palca artefaktów i regionalnych odsłonięć skalnych (Rys. uzupełniający 4 ) ujawnia, że homininy zajmowały się pozyskiwaniem do 12 km w basenie32 , 33 , a także eksploatowały kwarcyt lokalnie, 400 m na południe (Naisiusiu). Makroskopowo, cztery typy kwarcytu o odrębnym kolorze, wielkości ziarna, teksturze i składzie mineralnym były preferowanymi surowcami. Analiza tych odmian w odniesieniu do etapu redukcji (Rys. uzupełniający 2d ) wskazuje, że grupy oldowajskie celowały w określone typy kwarcytu dla płatków, podczas gdy inne warianty były przeznaczone na retuszowane narzędzia. Sferoidy i siekacze były wykonane z kwarcytu i ignimbrytu, dostępnych z koryt rzek.
Wczesna ekologia Oldowanu około 2 mln lat temu
Najwcześniejsza obecność Oldowanów w Ewass Oldupa, datowana na 2,03 Ma, ma miejsce w krajobrazie po erupcji, występując na rozległym strumieniu ignimbrytowym 34 , który pogrzebał sieć hydrologiczną południowo-wschodniej części basenu 18, jednocześnie otulając metamorficzne ostańce na zachodzie (rys. 1a i 2a, b ). Początkowe użycie homininów, udokumentowane w postaci 10 kamiennych narzędzi, pojawia się 1 m nad ignimbrytem Naabi i 17 m poniżej Tuff IA. Miało to miejsce po stabilizacji krajobrazu w powiązaniu z krętą, meandrującą rzeką płynącą na północny zachód, jak ujawniono w wykopie nr 7: ignimbryt Naabi był wynikiem wysokoenergetycznej, katastrofalnej erupcji wulkanicznej i towarzyszącego jej przepływu gruzu, który drastycznie zmienił krajobraz. Po tym wydarzenia wulkaniczne miały mniejszy wpływ i mogły rozwinąć się stabilne środowiska, takie jak koryta rzeczne i równiny zalewowe. W środowisku paleogeograficznym, w którym podłoże kwarcytowe wyłaniało się z wyrzuconego piroklastycznego strumienia, a kanał słodkiej wody przebiegał przez dystalną część stożka wulkanicznego, homininy okresowo wykorzystywały podnóże ostańca, w niewielkiej odległości od obu materiałów do produkcji narzędzi, a także źródeł wody z równiny zalewowej. Kontekst kwiatowy został ustalony przez fitolity (ryc. 5a , 6 i ryc. uzupełniająca 5 ), w których kilka typów paproci35 , 36 , 37 dominuje w zespołach (ryc. 6a, d, e ), co sugeruje istnienie łąki paprociowej z niewielkim wzrostem drzewiastym i trawami; w ten sposób pionierskie paprocie paproci ułatwiły ponowne ustanowienie zbiorowisk drzewiastych i trawiastych po spadku niszczycielskich zdarzeń wulkanicznych38 .
Ryc. 5: Geometria odsłonięcia, architektura stratygraficzna i idealna roślinność w Ewass Oldupa.

Lokalizacja zmierzonych przekrojów stratygraficznych i wykopów. Szczegóły przekroju 63-1-L i 63-1 przedstawiono na rys. 1b . Wizja artystyczna zbiorowisk roślinnych na przestrzeni czasu: a Łąka paprociowa po erupcji. b Mozaiki CFCT. c Las z palmami i paprociami. d Tereny trawiaste współczesne Tuff IA. e Otwarty las. f Zarośla z przewagą astrowatych.
Obraz w pełnym rozmiarze
Ryc. 6: Mikrofotografie wybranych fitolitów i pyłku z złoża I.
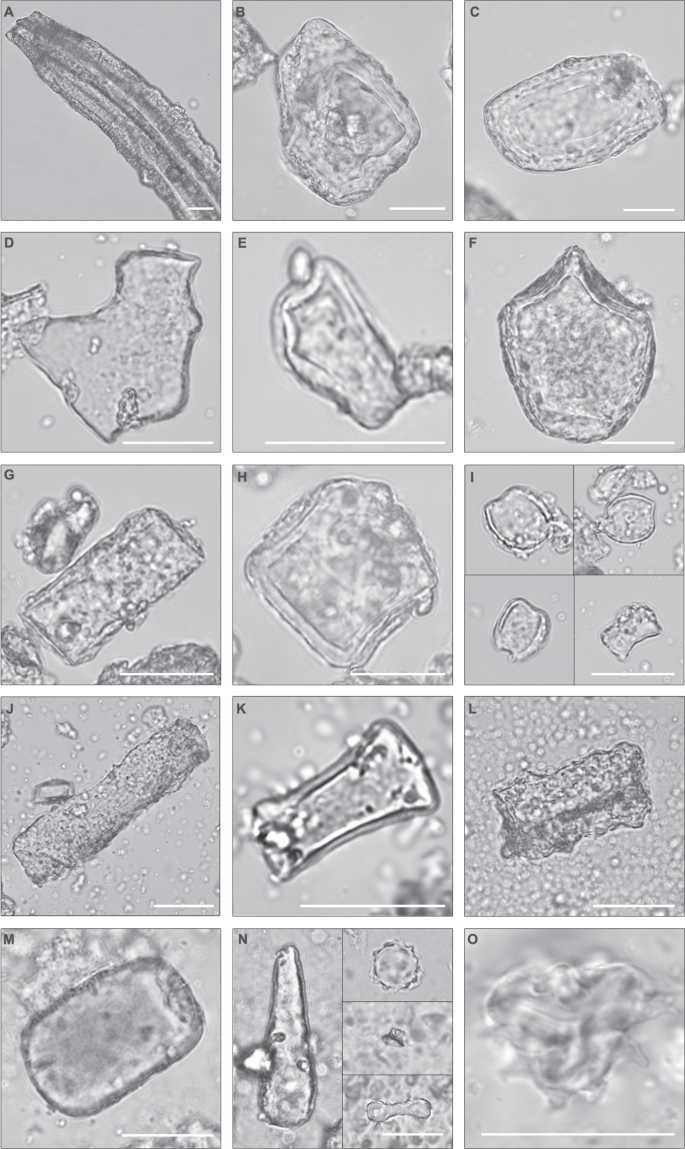
a Duży, tabliczkowy bruzdkowany cf. Pteridaceae. b , c Bryłowate fitolity z drzewiastych roślin dwuliściennych. d Część naskórka cf. Pteridaceae. e Tabularny rozdwojony cf. Pteridaceae (np. Cyrtomium ). f Bulliform, Poaceae. g Tabularny. h Tarczowaty cf. Solanaceae. i Siodłowy, krótki, Chloridoideae. j Tabularny skrobulowany. k Tabularny zduszczony cf. Salvadora . l Tabularny zatokowaty z drzewiastych roślin dwuliściennych. m Blokowaty z drzewiastych roślin dwuliściennych. n Po lewej. Zgrubiały z drzewiastych roślin dwuliściennych. Po prawej. Górny: kulisty jeżowaty, Arecaceae. Środkowy: wieżowy, Poaceae. Dolny: dwupłatkowy, długi, wypukły, Poaceae. o Pyłek astrowatych z górnego złoża I, bezpośrednio pod tufem Ng’eju. Stężenie pyłku = 2649; C3 Ret. Lamiaceae/Convolvulaceae, 101; Asteraceae, 72; Rutaceae, 2. Wszystkie paski skali = 25 µm. .
Obraz w pełnym rozmiarze
Podczas osadzania CFCTcz (2,015 ± 0,006 Ma) 25 Ma, ryzolity w jednostkach drobnoziarnistych reprezentują paleozole zalewowe równiny rzecznej w wykopie nr 5 (rys. 1 , 2c i 5 ). Przepływy masowe nie pozwoliły na przedłużony okres spoczynku, a zalewanie laharyczne pogrzebało synchroniczne krajobrazy leśne i pozostałości oldowańskie składające się z 12 kamieni wykopanych 1 km na zachód przez inne zespoły39 . Mikrowęgle drzewne z pożarów lasów osiągnęły szczyt (16 830 cząstek/cm3 ) (zakres w sekwencji stratygraficznej = 72–16 830/cm3 , mediana = 418/cm3 ) jednocześnie ze wzrostem opalu trawiastego (rys. uzupełniające 5a, b ). Duża różnorodność morfotypów fitolitów wskazuje na wyraźnie nierównomierne społeczności i szybkie zmiany w dominacji roślin (Rys. uzupełniający 5d, e ).
Trzy dodatkowe zespoły oldowańskie (~2 Ma) znaleziono na styku formacji Ngorongoro i złoża I, w powiązaniu z krętym kanałem płynącym na wschód i zachowane 4,6 m poniżej Tuff IA w wykopie nr 5 (rys. 1 , 2f, g , 3 i 5 ). Zrekonstruowane środowisko jest krótką mozaiką leśną, na co wskazują liczne dane zastępcze. Na przykład izotopy wosku roślinnego, przy użyciu średniej ważonej δ 13 C C 27 –C 33 dla każdej indywidualnej próbki i naniesione przy użyciu modelu mieszania sinus-kwadrat z wartościami końcowymi −30‰ (czysty C 3 ) i −19‰ (czysty C 4 ), ujawniają kontekst depozycyjny mozaik (rys. 7a ). Jedna próbka ilustruje środowisko zdominowane przez C3 prawdopodobnie w odpowiedzi na zwiększone opady deszczu (δ 13 C: −27,9‰), podczas gdy inne próbki wykazują wartości δ 13 C bardziej wskazujące na mieszane środowiska, wahające się od δ 13 C −22,6‰ do δ 13 C −24,6‰. Ten rozkład δ 13 C dla Ewass Oldupa jest analogiczny do dzisiejszych lasów akacji wschodnioafrykańskiej – Commiphora . Innym dowodem wskazującym na mozaikę leśną są stabilne izotopy ze szkliwa zębów: cały analizowany zespół reprezentuje czasy sprzed Tuff IA (δ 13 C: −7,2‰ do 1,6‰), co wskazuje na roślinożerców spożywających mieszankę roślin C3 i C4 ( ryc. 8 ). Chociaż pomiary δ 18 O mogą być pod wpływem szeregu czynników, w tym opadów, temperatury, źródła opadów i fizjologii, uzyskany zakres (δ 18 O: −4,7‰ do −0,2‰) jest zgodny ze zwierzętami pobierającymi wodę z podobnego źródła i nie ma wyraźnych rozróżnień między taksonami (ryc. 8 , ryc. uzupełniająca 6 i tabela uzupełniająca 3 ). Zakresu δ 18 O nie można było rozdzielić na obligatoryjny lub nie obligatoryjny status picia. Profil izotopowy szkliwa z Ewass Oldupa, w najniższym złożu I, przypomina profil z młodszych lasów w złożu II (ryc. 8 ), ale kontrastuje z otwartym profilem trawiastym przeważającym w penewspółczesnych lokalizacjach w zachodniej Kenii. Dodatkowym dowodem potwierdzającym rekonstrukcję paleośrodowiska mozaiki leśnej/trawiastej są fitolity (Tabela 2 , Ryc. 5 , 6 i Ryc. uzupełniający 5b ), charakteryzujące się zmienną produkcją zdrewniałych roślin dwuliściennych i traw.

a Rekonstrukcja krajobrazu roślinnego przy użyciu δ 13 C średniej ważonej n- alkanów C 27 – C 33. Model mieszania sinus-kwadrat z wartościami końcowymi −30‰ (dla czystego C 3 ) i −19‰ (dla czystego C 4 ) został użyty do wizualizacji ekologii roślin podczas osadzania próbki. δ 13 C n -alkanów zmienia się między −29‰ i −39‰ w istniejących roślinach C 3 oraz między −14‰ i −26‰ w roślinności C 4 . VPDB Vienna Pee-Dee Belemnite. b – f Wykresy pokazują względną obfitość każdego związku n -alkanowego (szare słupki), ich wartości δ 13 C (czarne kółka) i średnią długość łańcucha (ACL) homologu węglowego C 25 – C 35 . Cyfry rzymskie oznaczają pochodzenie w kolumnie stratygraficznej od dołu do góry, a są to pojedyncze próbki, a nie kompozyty. Umiejscowienie każdej próbki wzdłuż gradientu gleby/rośliny opiera się na uzyskanej średniej ważonej dla każdej próbki.
Obraz w pełnym rozmiarze
Ryc. 8: Pomiary stabilnej zawartości węgla (δ13C) i tlenu (δ18O) w zębach zwierząt z Ewass Oldupa (poniżej Tuff IA) w porównaniu z ówczesnymi i młodszymi zbiorami danych kopalnych 14 , 75 .

Test Manna–Whitneya–Wilcoxona wykazuje istotną różnicę w δ13C [ W = 215, <0,05(0,004)]. Istnieje również rozróżnienie δ18O: [ W = 92, <0,05(0,000)]. Ogólnie rzecz biorąc, wyniki sugerują, że okres 2,0–1,9 Ma był wilgotniejszy niż 1,8–1,6 Ma. Uwaga: w porównaniu uwzględniono tylko dane z tych samych rodzin taksonów.
Obraz w pełnym rozmiarze
Piąty zespół oldowański obejmuje narzędzia kamienne i kości 4 m poniżej Tuff IA (~2 Ma) na transgresywnym brzegu jeziora, co stanowi najwcześniejszą i najdalej na zachód wysuniętą ekspansję paleojeziornego Oldupai, jak odsłonięto w wykopie nr 3 (ryc. 1 , 2e , 3 i 5 ). Zróżnicowana taksonomicznie fauna bez modyfikacji homininów obejmuje Antelopini, Bovini, Hippotragini i Tragelaphini (tabela uzupełniająca 4 , uwaga uzupełniająca 1 ), pokazując mieszane siedliska, oprócz taksonów zależnych od wody. Fitolity pokazują florystyczny tandem lasów i palm, który jest uosobieniem zajęć na brzegu jeziora oldowańskiego charakterystycznych dla Upper Bed I 40, 41, 42, 43 (ryc. 5–6, tabela 2 ) , co stanowi bliskość słodkiej wody i preferowanych źródeł surowców dla homininów. Ponadto narzędzia kamienne z kultury oldowajskiej z tego poziomu (ryc. 4e, f ) są najwcześniejszymi dowodami kamiennymi osadzonymi w woskowej glinie pochodzącej z słonego/zasadowego środowiska jeziornego.
Szósty klaster oldowański znajdował się w pobliżu meandrującego strumienia w wielopiętrowym pasie kanałów opadających do jeziora, położony 3 m pod Tuff IA (~2 Ma) i odkryty w wykopie nr 2 (rys. 1 , 2f , 3 i 5 ). Fauna nie wykazuje śladów przetwarzania przez homininy (Uwaga uzupełniająca 1 ). Obejmuje ona wołowate, suidy, koniowate, duże ssaki i plio-plejstoceńskie cercopithecine, które żerują w środowiskach trawiastych (por. Theropithecus oswaldi ) 44 , 45, a także wieloletnie wskaźniki wody ( Chelonia , Crocodylus i Hippopotamus ) (Tabela uzupełniająca 4 ). Fitolity pokazują cykliczny rozkwit i zanik zdrewniałych roślin dwuliściennych i traw46 , natomiast wkraczanie traw na tereny leśne wystąpiło dwukrotnie (Rys. uzupełniający 5b ).
Wartości izotopowych n -alkanów (C 29 C 31 C 33 ) dla Górnego Złoża I z regresywnych brzegów jezior i małych strumieni między Tuff IA-IB pokazują otwarte tereny leśne z mieszanymi siedliskami C 3 – C 4 (ryc. 1 i 7a, d–f ) w pozycji stratygraficznej równoważnej najstarszemu Homo habilis w basenie wschodnim 30 (OH 24). Sukcesja zajęć oldowańskich kończy się artefaktami z kontekstu brzegu jeziora umieszczonymi między Tuff IB i IC (1,848/1,832 Ma), w pozycji i środowisku sedymentacyjnym, które na zachodnim brzegu jeziora jest odpowiednikiem poziomu 22 Zinj 23 , 24 , 41 , 47 , podczas gdy nadal 2 m poniżej OH 65 21 (ryc. 1 ), jak odkryto w wykopie nr 6. Ziarna pyłku wyrażają bezdrzewny, beztrawny, hiperkseryczny step zdominowany przez astrowate w kohorcie z Lamiaceae/Convolvulaceae i zaroślami Rutaceae (ryc. 6o ) bezpośrednio pod tufem Ng’eju, korelującym z poziomem mułowcowo-wapiennym nad OH 65 22 , 48 .
Dyskusja
Ewass Oldupa to wczesnooldowańskie stanowisko warstwowe o wysokiej rozdzielczości, wieloetapowe, które poprzedza i rozciąga się nad Bed I. To stanowisko zawiera najstarsze dowody Oldupai na pozostałości kulturowe homininów związane z siedliskami z zaburzeń po wulkanicznych, a zatem wczesne przykłady adaptacji do głównych przekształceń geomorfologicznych i ekologicznych. Najwcześniejsze narzędzia oldowajskie pojawiają się bezpośrednio po osadzeniu ignimbrytu Naabi i są związane z ułożoną serią rzeczną złożoną z mafijnego tephry (ryc. 1 ). Tam, poprzecznie warstwowany piaskowiec i ogólne wznoszące się klarowanie przykryte mułowcem wskazują, że ludzie zaopatrywali się w pobliżu meandrujących rzek. Ta epizodyczna eksploatacja środowisk rzecznych poprzedza niestabilne powierzchnie charakterystyczne dla młodszych horyzontów CFCT 39 . Duże zespoły skalne i faunistyczne z kontaktu między najwyższym CFCT a najniższym Bed I, wraz z szeroką gamą zastępczych roślinności, oznaczają początek osadnictwa jeziornego w basenie, które poprzedza umiejscowienie Tuff IA. Zestawy narzędzi oldowańskich z tego okresu przecinają wcześniejsze i współczesne technologie z Etiopii i Kenii, jednocześnie rozwijając typy narzędzi, takie jak sferoid powszechnie spotykany w młodszych zespołach 24 . Jeśli te narzędzia kamienne napędzały zaopatrzenie w nieprzewidywalnych środowiskach, to za pośrednictwem strategii generalistycznej 49 , 50 dla różnych zadań, które nie obejmowały obdzierania z mięsa tusz zwierzęcych, jak pokazuje brak zmodyfikowanej przez człowieka fauny.
Podsumowując, najwcześniejsi hominini Oldupai zajmowali heterogeniczne, niestabilne środowiska. Dane sedymentacyjne, chemiczne i roślinne wskazują, że wczesni hominini Olduwai byli pionierami szybkiego zajmowania geo-ustawień, które przechodziły drastyczne zmiany w rozmieszczeniu i strukturze zasobów hydrologicznych oraz wspierały nierówną florę. Odkryte dowody wskazują na okresowe, powtarzające się użytkowanie ziemi w podzbiorze środowisk przerywanych okresami, w których nie ma dowodów na aktywność homininów. Nasza praca ujawnia wczesne narzędzia kamienne Olduwai w zróżnicowanych środowiskach fizycznych, z oznakami, że środowiska te zmieniały się znacząco w przestrzeni i czasie. Ponadto grupy Olduwai wykorzystywały środowiska zaburzeń — odkrycie, które jest unikalne dla tego okresu i przedstawia złożone zachowanie wśród wczesnoplejstoceńskich homininów. Zdolność do eksploatacji zróżnicowanych biomów umożliwiła homininom ekspansję poza Afrykę. Ta elastyczność behawioralna i zestaw zdolności adaptacyjnych urzeczywistniły się w sytuacji braku kontrolowanego ognia, w otoczeniu dużych drapieżników, i można je interpretować jako wskaźnik inwazyjności, który ułatwił wczesne globalne rozproszenie Homo.
Metody
Biomarkery
Suchy, zhomogenizowany osad ekstrahowano metodą PSE (Büchi Speed Extractor E-916) w trzech 10-minutowych cyklach w 9:1 (v:v) DCM:MeOH w temperaturze 100 °C i ciśnieniu 103 barów/1500 psi. Neutralne, niepolarne węglowodory oddzielono od całkowitych ekstraktów lipidowych metodą chromatografii kolumnowej aminopropylowej przy użyciu 2:1 DCM:IPOH. Następnie normalne ( n -) alkany oddzielono od frakcji neutralnej przy użyciu chromatografii kolumnowej na żelu krzemionkowym z dodatkiem azotanu srebra przy użyciu heksanu. Analizę chromatografii gazowej (GC) wykonano przy użyciu 7890B (Agilent) i kolumny HP-5 (Agilent; długość 30 m, średnica wewnętrzna 0,25 mm i film 0,25 µm) z 5977 A MSD. Próbki wstrzykiwano w temperaturze 250 °C w trybie splitless, a temperaturę pieca zaprogramowano od 60 °C (utrzymanie 1 min) do 150 °C z szybkością 10 °C/minutę, a następnie do 320 °C z szybkością 6 °C/minutę (utrzymanie 10 min). Źródło spektrometrii masowej (MS) działało w temperaturze 230 °C przy 70 eV i pełnej szybkości skanowania m/z 50–550. Woski roślinne n- alkany identyfikowano poprzez porównanie widm masowych i czasu retencji z zewnętrzną mieszaniną standardową (C 21 –C 40 ).
Izotop-stosunek MS Wartości δ 13 C specyficzne dla związku analizowano za pomocą GC 7890B (Agilent) z kolumną kapilarną HP-5 (Agilent) obsługiwaną w trybie rozdzielonym sprzężoną ze spektrometrem masowym Elementar Isoprime AnthrovisION za pomocą interfejsu systemu pieca Elementar GC5 pracującego w temperaturze 850 °C. Temperaturę pieca zaprogramowano od 60 °C (przytrzymanie 1 min) do 150 °C przy 10 °C/minutę, a następnie do 320 °C przy 6 °C/minutę (przytrzymanie 10 min). Gazem nośnym był hel ze stałym przepływem 1,1 ml/min. Dokładność wartości δ 13 C oceniano w odniesieniu do międzynarodowej standardowej mieszanki laboratoryjnej (Indiana B4, Arndt Schimmelmann, University of Indiana) wstrzykiwanej na początku i na końcu każdego zestawu próbek. Odchylenie standardowe roboczego wzorca n -alkanowego wynosiło ≤0,5‰. Stosunek izotopów jest wyrażony jako wartości δ 13 C w promilach w stosunku do wzorca Vienna Pee Dee Belemnite (VPDB).
Fluorescencja rentgenowska z dyspersją energii
Artefakty kwarcytowe ( n = 24) poddano indywidualnej sonikacji (10 min; 40 kHz), a następnie charakteryzowano chemicznie przy użyciu EDXRF (Thermo Scientific ARL QUANT’X; promieniowanie hamowania okna końcowego; lampa rentgenowska Rh target 50 W; okno berylowe 76 μm; 4–50 kV; pompa próżniowa Edwards RV8). Każdy cykl analityczny ( n = 5) obejmował standard odniesienia USGS RGM-2 do kalibracji przyrządu. Docelowe pierwiastki obejmowały osiem głównych tlenków (SiO 2 , TiO 2 , Al 2 O 3 , Fe 2 O 3 , MnO, MgO, CaO i K 2 O) i 10 pierwiastków śladowych (Cu, Zn, Rb, Sr, Y, Zr, Nb, Ba, Pb i Th). Intensywności promieniowania rentgenowskiego zostały automatycznie przeliczone na szacunki stężeń (główne tlenki = % wag.; pierwiastki śladowe = ppm) przy użyciu linii kalibracji najmniejszych kwadratów ilorazowej do rozrzutu Comptona dla każdego pierwiastka w oparciu o międzynarodowe wzorce odniesienia (AGV-2, BCR-2, BHVO-2, BIR-1a, GSP-2, JR-1, JR-2, QLO-1, RGM-2, SDC-1, STM-2, TLM-1 i W-2a). Szacunki stężeń zostały znormalizowane zgodnie ze standardem odniesienia USGS RGM-2 przy użyciu skalowania Min–Max, przy czym każda wartość została przeskalowana do zakresu [0, 1]. Te same warunki analityczne i metody obróbki danych zostały wdrożone dla próbek geologicznych kwarcytu ( n = 125) z pięciu odsłonięć, które stanowią zbiór odniesienia stosowany do analiz porównawczych51 , 52 . Eksplorację danych i liniową analizę dyskryminacyjną znormalizowanych danych chemicznych przeprowadzono w programie RStudio przy użyciu 14 pakietów 51 , 52 .
Wykop
Lokalizację materiałów rejestrowano na Leica TS 09 podczas wykopalisk. Materiały umieszczano w układzie kartezjańskim X – Y , a wartości Z rejestrowano również przed indywidualnym etykietowaniem i pakowaniem. Osady z każdego wykopu przesiewano na sucho (oczka 0,5 cm). Identyfikację i współrzędne XYZ eksportowano do bazy danych. Ta baza danych terenowych stanowiła macierz dla klasyfikacji techno-typologicznej, taksonomii, tafonomii i analizy przestrzennej. Dane zarejestrowane w 3D zostały naniesione na wykres XY przy użyciu ArcMap10.7, a projekcje XZ i YZ wykonano w kierunkach N–S i E–W. Do danych XY zastosowano analizę gęstości jądra (promień wyszukiwania = 25 cm) .
Fauna
Badano elementy >2 cm. W celu identyfikacji taksonomicznej skonsultowaliśmy cyfrowe archiwum ssaków z Afryki Wschodniej Smithsonian Institution 53 , podręcznik osteologiczny 54 , ustaloną kolekcję referencyjną muzeum dostępną bezpośrednio dla nas w IPHES (Tarragona, Hiszpania), a także specjalistyczną literaturę faunistyczną na temat skamieniałości z wczesnego plejstocenu 55 . Modyfikacje powierzchni odnotowano dla wszystkich okazów z korą. Następnie okazy badano pod stereomikroskopem (Optech HZ, 10–60x). Profile szkieletowe utworzono przy użyciu wartości liczby zidentyfikowanych okazów (NISP). Klasyfikacja klas wielkości 56 , 57 , 58 została zastosowana tam, gdzie nie można było dokonać klasyfikacji wyższego rzędu. Minimalną liczbę elementów (MNE) obliczono na podstawie NISP, odnotowując częstość, części, rozmiar i stronę podlegającą analizie 59 . Części MNE wykorzystano do standaryzacji minimalnych jednostek zwierzęcych (% MAU) 60 . Współczynnik korelacji dla procentu MAU i gęstości mineralnej kości 61 , biorąc pod uwagę MNE każdej części wszystkich pierwiastków wśród kopytnych. Wiek okazu podano, jeśli to możliwe, w przeciwnym razie użyto „nieznany”. Zanotowano wzory złamań dla kości długich i płaskich 62 .
Geochemia mineralna
Skład fenokryształów określono za pomocą analizy mikrosondy elektronowej. Wstępna analiza petrograficzna polerowanych cienkich przekrojów została przeprowadzona przy użyciu mikroskopu petrograficznego z polaryzacją krzyżową w celu określenia liczebności fenokryształów, sortowania, stopnia zmian i przydatności do analizy. Próbki pokryto węglem i przeanalizowano za pomocą mikrosondy elektronowej JEOL JXA-8200. Wiązka mikrosondy elektronowej była obsługiwana przy natężeniu prądu 8 μA i napięciu 15 kV oraz średnicy wiązki 11 mm. Do identyfikacji potencjalnych ziaren minerałów do analizy wykorzystano kombinację jasności z obrazów BSE i EDS. Tam, gdzie było to możliwe, analizowano co najmniej 10 ziaren na próbkę na minerał (30 punktów/próbkę). Standardami kalibracji mikroprobówki dla ziaren skalenia były ortoklaz i albit, standardami dla ziaren augitu/klinopiroksenu były Cr-augit i hornblenda, a ilmenit został użyty do kalibracji dla ziaren tlenkowych. Podjęto wysiłki, aby przeanalizować ziarna zawarte w szkle lub w materiale pumeksowym. Ziarna mineralne analizowano losowo, aby zapobiec stronniczości. Całkowite procenty wagowe dla analiz skalenia i piroksenu mniejsze niż 96% lub większe niż 102% zostały wyłączone z ostatecznych danych, podobnie jak ziarna tytanomagnetytu o łącznych wartościach mniejszych niż 90%.
Analiza fitolitów
Próbki (3 g) zmieszano z 0,1% heksametafosforanem sodu (NaPO3 ) 6 i poddano działaniu ultradźwięków (5 min). Wytrząsanie orbitalne prowadzono przez noc (200 obr./min). Po rozproszeniu glinki zastosowano 3 N kwas solny (HCl) i kwas azotowy (HNO3) oraz nadtlenek wodoru (H2O2). Poliwolframian sodu (3Na2WO4 · 9WO3 · H2O ) ( Poly – Gee) o ciężarze właściwym 2,4 oddzielił fitolity. Płukanie i wirowanie wykonywano przy 3000 obr./min przez 5 min . Aliquot ( 0,001 g ) umieszczono na szkiełku mikroskopowym z Entellan New (pokrycie: 20 × 40 mm = obszar badany). Mikroskopia systemu wykonywano przy powiększeniu 40x (Olympus BX41, Motic BA410E). W miarę możliwości liczono co najmniej 200 fitolitów lub do momentu wyginięcia próbki. Referencyjne dane obejmowały kilka afrykańskich ekoregionów 46 , 63 , 64 , 65 , 66 , 67 , 68 oraz wcześniejsze prace w wąwozie Oldupai 43 , 69 . Próbki archeologiczne porównano z bazą 29 gatunków roślin i 35 gleb z mozaik leśnych Acacia – Commiphora z badanego obszaru, stosując ogólne i diagnostyczne podejścia do rekonstrukcji fizjonomii i rangi pokrywy roślinnej poprzez sygnał ich fitolitów 46 . Nomenklatura klasyfikacji była zgodna z ref. 70 .
Pyłek i mikrowęgiel
Protokół został zaadaptowany z ref. 71. Próbkę (2 ml) zmieszano z 3% heksametafosforanem sodu (NaPO3 ) 6 w łaźni wodnej (90 °C) przez 30 min i mieszano mechanicznie przez kilka dni. Dodano tabele zarodników Lycopodium w celu obliczenia stężeń pyłku i mikrowęgla. Próbki osiadły i zdekantowano (<2 µm), a następnie przesiano (>125 µm). Oddzielenie pyłku i węgla drzewnego przeprowadzono za pomocą delikatnego mieszania mechanicznego przez kilka dni, a następnie przeprowadzono separację ciężkiej cieczy przy użyciu heteropoliwolframianu litu o ciężarze właściwym 2,0. Acetolizę uzyskano poprzez mieszaninę 9:1 C 4 H 6 O 3 do H 2 SO 4 i 10% HCl. Próbki umieszczono na szkiełku mikroskopowym z gliceryną (pokrywa: 22 × 40 mm = obszar sprawdzany). Mikroskopię systemową wykonano przy powiększeniu 25x, 40x, 63x. Zliczono co najmniej 350 ziaren pyłku i 200 mikrowęgli.
Analiza stabilnych izotopów węgla i tlenu w szkliwie zębów zwierząt
Wszystkie wybrane okazy miały doskonałe warunki konserwacji, nie uwzględniono żadnych zwietrzałych lub pofragmentowanych okazów dentystycznych. Zęby oczyszczono za pomocą abrazji powietrznej. Próbkę proszku szkliwa (6 mg) pobrano z krawędzi policzkowej za pomocą wiertła diamentowego. Płukano ją w 1,5% NaClO przez 60 min, płukano, a następnie odwirowywano. Następnie płukano ją kwasem octowym (0,1 M) przez 10 min i płukano. Pozostałość liofilizowano przez 24 h, a następnie poddano reakcji z H3PO4 , a gazy mierzono pod kątem stabilnych stosunków węgla i tlenu za pomocą GasBench II (Thermo) połączonego z Delta V Advantage MS (Thermo). Wartości porównano z normami międzynarodowymi: IAEA-603 (δ 13 C = 2,5; δ 18 O = −2,4); IAEA-CO-8 (δ 13 C = −5,8; δ 18 O = −22,7); USGS44 (δ 13 C = −42,2) i wewnętrzny standard MERCK (δ 13 C = −41,3; δ 18 O = −14,4). Błąd pomiaru standardu MERCK c wynosił ±0,1‰ dla δ 13 C i ±0,2‰ dla δ 18 O. Całkowita precyzja pomiarów poprzez powtarzane ekstrakty z wewnętrznego standardu szkliwa zębów bydlęcych c wynosiła ±0,2‰ dla δ 13 C i ±0,3‰. Obowiązkowy lub nieobowiązkowy status picia został wywnioskowany z zakresu δ 18 O zgodnie z odniesieniami 72 , 73 , 74 .
Narzędzia kamienne
Wszystkie okazy zostały zmierzone i zważone, a atrybuty zarejestrowane w bazie danych. Rozróżniono klasy technologiczne, takie jak rdzeń, fragment rdzenia, odłupek, połamany odłupek, fragment odłupka z platformą, materiał perkusyjny, kawałek retuszowany i narzędzia. Metody kucia sklasyfikowano według facelności (zwracając uwagę na liczbę eksploatowanych powierzchni) i polarności (kierunek eksploatacji z platformy uderzeniowej). Zaobserwowano następujące metody redukcji rdzenia: (i) wielobiegunowo-wielopłaszczyznowa: rdzenie z czterema lub więcej powierzchniami eksploatowanymi w co najmniej czterech różnych kierunkach; (ii) jednobiegunowo-podłużna: rdzenie z jedną powierzchnią eksploatowaną jednokierunkowo; (iii) ortogonalnie-dwupłaszczyznowa: rdzenie z dwiema przeciwległymi powierzchniami kutymi jednokierunkowo; (iv) dośrodkowo-dwupłaszczyznowa: rdzenie z dwiema powierzchniami eksploatacyjnymi zorganizowanymi przez płaszczyznę poziomą, każda strona z co najmniej trzema usunięciami dośrodkowymi; (v) centripetal-unifacial: rdzenie z jedną powierzchnią eksploatowaną i co najmniej trzema ekstrakcjami dośrodkowymi; (vi) bipolar-on-anvil: rdzenie z dowodami umieszczenia na pasywnej powierzchni; (vii) bipolar-unifacial: rdzenie z jedną powierzchnią eksploatowaną dwukierunkowo. Wielowymiarowa ordynacja wykorzystała zmienne z ref. 15 poprzez oprogramowanie Palaeotological Statistics . Zmienne zostały znormalizowane. Pierwsze trzy komponenty PCA wyjaśniają 80,1% wariancji.
Podsumowanie raportu
Więcej informacji na temat projektu badawczego można znaleźć w podsumowaniu badań Nature Research Reporting Summary dostępnym pod tym artykułem.
Dostępność danych
Wszystkie dane są dostępne w tekście głównym i materiałach uzupełniających. Materiały mają być zdeponowane w Narodowych Muzeach Tanzanii.
Odniesienia
-
Toth, N. & Schick, K. Przegląd implikacji poznawczych kompleksu przemysłowego Oldowan. Azania 53 , 3–39 (2018).
-
Pobiner, BL, Rogers, MJ, Monahan, CM i Harris, JW Nowe dowody na strategie przetwarzania tuszy homininów po 1,5 mln lat, Koobi Fora, Kenia. J. Hum. Evol. 55 , 103–130 (2008).
-
Domínguez Rodrigo, M. Spożywanie mięsa przez wczesne hominidy na stanowisku FLK 22 Zinjanthropus, wąwóz Olduvai, Tanzania: podejście eksperymentalne z wykorzystaniem danych z śladów nacięć. J. Hum. Evol. 33 , 669–690 (1997).
-
Sahnouni, M. i in. Artefakty sprzed 1,9 i 2,4 miliona lat oraz kości z nacięciami kamiennymi z Ain Boucherit w Algierii. Science 362 , 1297–1301 (2018).
-
Ferraro, JV i in. Najwcześniejsze dowody archeologiczne na przetrwałą mięsożerność homininów. PLoS One 8 , e62174 (2013).
-
Braun, DR i in. Wczesna dieta homininów obejmowała różnorodne zwierzęta lądowe i wodne 1,95 MA w East Turkana, Kenia. Proc. Natl Acad. Sci. 107 , 10002–10007 (2010).
-
Zhu, Z. i in. Osiedlenie chińskiego płaskowyżu lessowego przez hominidy od około 2,1 mln lat temu. Nature 559 , 608–612 (2018).
-
Lordkipanidze, D. i in. Kompletna czaszka z Dmanisi w Gruzji i biologia ewolucyjna wczesnego Homo. Sci. 342 , 326–331 (2013).
-
Ferring, R. i in. Najwcześniejsze ślady osadnictwa ludzkiego w Dmanisi (Gruziński Kaukaz) datowane na 1,85–1,78 mln lat temu. Proc. Natl Acad. Sci. 108 , 10432–10436 (2011).
-
Asfaw, B. i in. Fejej: nowy obszar badań paleoantropologicznych w Etiopii. J. Hum. Ewolucja 21 , 137–143 (1991).
-
Barksy, D. i in. Wczesnoolduwajski zespół narzędzi kamiennych z Fejej FJ-1A. Etiopia. J.Afr. Archeol. 9 , 207–224 (2011).
-
Kimbel, W. i in. Późnoplioceńskie narzędzia Homo i Olduwajskie z formacji Hadar (członek Kada Hadar), Etiopia. J. Hum. Ewolucja 31 , 549–561 (1996).
-
Patterson, DB i in. Ewolucja ekosystemu i paleobiologia homininów w East Turkana, północna Kenia, między 2,0 a 1,4 mln lat temu. Palaeogeogr. Palaeoclimatol. Palaeoecol. 481 , 1–13 (2017).
-
Plummer, TW i in. Najstarsze dowody na istnienie homininów wytwarzających narzędzia w ekosystemie zdominowanym przez tereny trawiaste. PLoS One 4 , e7199 (2009).
-
Braun, DR i in. Najwcześniejsze znane artefakty oldowańskie sprzed >2,58 Ma z Ledi-Geraru w Etiopii podkreślają wczesną różnorodność technologiczną. Proc. Natl Acad. Sci. 116 , 11712–11717 (2019).
-
Plummer, T. Łupane kamienie i stare kości: ewolucja biologiczna i kulturowa u zarania technologii. Yearb. Phys. Anthropol. 47 , 118–164 (2004).
-
Plummer, TW i Bishop, LC Zachowanie i ekologia człowiekowatych oldowańskich w Kanjera South w Kenii. J. Anthropol. Sci. 94 , 1–12 (2016).
-
Stanistreet, IG i in. Nowa stratygrafia basenu Olduvai i koncepcje stratygraficzne ujawnione przez rdzenie OGCP w depocentrum Palaeolake Olduvai w Tanzanii. Palaeogeogr. Palaeoclimatol. Palaeoecol. tom 554 (2020).
-
Bobe, R. i in. Zmiany fauny, zmienność środowiskowa i ewolucja człowiekowatych w późnym pliocenie. J. Hum. Evol. 42 , 475–497 (2002).
-
Harrison, T. Paleontologia i geologia Laetoli: Ewolucja człowieka w kontekście tom 1 (2011). Springer.
-
Plummer, TW i Finestone, E. W Rethinking Human Evolution (red. Schwartz, J.) (MIT Press, 2017). Rozdział 13.
-
Blumenschine, RJ i in. Późnoplioceńskie użytkowanie ziemi przez Homo i hominidy w zachodniej części wąwozu Olduvai w Tanzanii. Science 299 , 1217–1221 (2003).
-
Hay, R. Geologia wąwozu Olduvai (Univ. of California Press, 1976).
-
Leakey, MD Wąwóz Olduvai: wykopaliska w warstwach I i II, 1960 – 1963 , tom 3 (Cambridge Univ. Press, 1971).
-
Deino, AL Datowanie 40 Ar/ 39 Ar w Bed I, wąwóz Olduvai w Tanzanii i chronologia zmian klimatycznych we wczesnym plejstocenie. J. Hum. Evol. 63 , 251–273 (2012).
-
McHenry, LJ Zmieniona struktura stratygraficzna dla złoża Olduvai Gorge Bed I oparta na geochemii tufu. J. Hum. Evol. 63 , 284–299 (2012).
-
McHenry, LJ i in. Odcisk palca tufu i korelacje między rdzeniami OGCP i odsłonięciami dla Pre-Bed I i warstw I/II w wąwozie Olduvai w Tanzanii. Palaeogeogr. Palaeoclimatol. Palaeoecol. 548 , 109630 (2020).
-
Habermann, JM i in. Dyskryminacja, korelacja i pochodzenie markerów tefrostratygraficznych Bed I, wąwóz Olduvai, Tanzania, na podstawie wieloczynnikowych analiz składu fenokryształów. Sediment. Geol. 339 , 115–133 (2016).
-
Stanistreet, IG i in. Paleosalinność i paleoklimatyczne wskaźniki geochemiczne (pierwiastki Ti, Mg, Al) zmieniają się wraz z cyklicznością Milankovitcha (1,3 do 2,0 Ma), rdzenie OGCP, jezioro Palaeolake Olduvai, Tanzania. Palaeogeogr. Palaeoclimatol. Palaeoecol. 546 , 109656 (2020).
-
Tobias, PV Olduvai Gorge: Czaszki, odlewy wewnętrzne i zęby Homo habilis , tom 4 (Cambridge Univ. Press, 1991).
-
Stanistreet, IG i in. Lahar zalał, zmodyfikował i zachował stanowisko Olduvai DK z wczesnymi hominidami (OH24 i OH56) sprzed 1,88 mln lat. J. Hum. Evol. 116 , 27–42 (2018).
-
Schick, K. & Toth, N. (red.) Oldowan: Studium przypadku wczesnej epoki kamienia łupanego (Stone Age Institute Press, 2006).
-
Reti, JS Kwantyfikacja produkcji narzędzi z kamienia oldowańskiego w wąwozie Olduvai w Tanzanii. PLoS One 11 , e0147352 (2016).
-
Mollel, GF, Swisher, CC III, Feigenson, MD i Carr, MJ Ewolucja geochemiczna kaldery Ngorongoro, Północna Tanzania: implikacje dla interakcji skorupa-magma. Planeta Ziemia. Nauka. Łotysz. 271 , 337–347 (2008).
-
Kondo, R., Childs, CW, Atkinson, IAE i Pritchard, T. Opal Phytoliths z Nowej Zelandii . (Manaaki Whenua Press, 1994).
-
Sundue, M. Ciała krzemionkowe i ich systematyczne implikacje w Pteridaceae (Pteridophyta). Bot . J. Linn. Soc. 161 , 422–435 (2009).
-
Chauhan, DK, Tripathi, DK, Since, P. & Tiwari, SP Biogeniczna krzemionka w niektórych paprotnikach. Bionature 29 , 1–9 (2009).
-
Walker, LR i Sharpe, JM w ekologii paproci (red. Mehltreter, K., Walker, LR i Sharpe, JM) (Cambridge Univ. Press, 2010). Rozdział 6.
-
Habermann, JM i in. Drzewa in situ ~2,0 mln lat temu odkryte jako skamieniałe pnie z korzeniami, najniższe złoże I, wąwóz Olduvai, Tanzania. J. Hum. Evol. 90 , 74–87 (2016).
-
Albert, RM, Bamford, MK i Esteban, I. Rekonstrukcja starożytnych krajobrazów roślinności palmowej przy użyciu podejścia fitolitycznego. Quat. Int. 369 , 1–16 (2014).
-
Uribelarrea, D. i in. Geoarcheologiczna i geometrycznie poprawiona rekonstrukcja paleokrajobrazu FLK Zinj z okresu 1,84 mln lat temu w wąwozie Olduvai w Tanzanii. Quat. Int. 322–323 , 7–31 (2014).
-
Magill, CR, Ashley, GM, Domínguez-Rodrigo, M. i Freeman, KH Opcje dietetyczne i zachowania sugerowane przez dowody biomarkerów roślinnych we wczesnym środowisku ludzkim. Proc. Natl Acad. Sci. 113 , 2874–2879 (2016).
-
Ashley, GM i in. Rekonstrukcja paleośrodowiskowa i paleoekologiczna oazy słodkowodnej na sawannie w FLK North, wąwóz Olduvai, Tanzania. Quat. Res. 74 , 333–343 (2010).
-
Jablonski, NG i Leakey, MG Projekt badawczy Koobi Fora: Skamieniałe małpy , tom 6 (California Academy of Sciences, 2008).
-
Jolly, CJ Klasyfikacja i historia naturalna Theropithecus ( Simopithecus ) (Andrews, 1916), pawianów z afrykańskiego pliocenu i plejstocenu. Bull. Br. Mus. Nat. Hist. Geol. 22 , 1–123 (1972).
-
Mercader, J. i in. Gleba i fitolity roślinne z mozaik Acacia-Commiphora w wąwozie Oldupai (Tanzania). PeerJ 7 , e8211 (2019).
-
Leakey, LSB Nowa skamielina z Olduvai. Nature 184 , 491–494 (1959).
-
Sikes, NE i Ashley, GM Stabilne izotopy węglanów pedogenicznych jako wskaźniki paleoekologii w plio-plejstocenie (górne złoże I), zachodni brzeg Kotliny Olduvai, Tanzania. J. Hum. Evol. 53 , 574–594 (2007).
-
Potts, R. Zmienne kontra modele użytkowania ziemi przez wczesnoplejstoceńskie homininy. J. Hum. Evol. 27 , 7–24 (1994).
-
Potts, R. Ewolucja homininów w warunkach dużej zmienności środowiskowej. Quat. Sci. Rev. 73 , 1–13 (2013).
-
Soto, M. i in. Odcisk palca odsłonięć kwarcytowych w wąwozie Oldupai. Tanzan. J. Archaeol. Sci. Rep. 29 , 102010 (2020).
-
Soto, M. i in. Systematyczne pobieranie próbek kwarcytu w analizie źródeł: zmienność wewnątrz odsłonięcia w Naibor Soit, Tanzania (część I). Archaeol. Anthropol. Sci. 12 , 1–14 (2020).
-
Assefa, Z Cyfrowe archiwum ssaków wschodnioafrykańskich . (Smithsonian Institution, Waszyngton, 2006).
-
Pales, L. i Lamert, P. Atlas osteóologique: pour servir à l’identification des mammifères du Quaternaire. Les Members Roślinożercy . (Centre National de la Recherche Scientifique, 1971).
-
Jablonski, NG i Leakey, MG Koobi w ramach projektu badawczego: The Fossil Monkeys Vol. 6 (California Academy of Sciences, 2008).
-
Brain, CK Niektóre proponowane procedury w analizie nagromadzeń kostnych z miejsc czwartorzędowych w południowej Afryce. Ann. Transv. Mus. 29 , 1–5 (1974).
-
Brain, CK Myśliwi czy ścigani? Wprowadzenie do afrykańskiej tafonomii jaskiniowej (Univ. of Chicago Press, 1981).
-
Klein, RG Fauna ssaków miejsc ujścia rzeki Klasies, Prowincja Przylądkowa, Afryka Południowa. S. Afr. Archaeol. Bull. 31 , 75–98 (1976).
-
Klein, RG i Cruz-Uribe, K. Analiza kości zwierzęcych ze stanowisk archeologicznych (Univ. of Chicago Press, 1984).
-
Binford, LR Pozostałości fauny z ujścia rzeki Klasies . (Academic Press, 1984).
-
Lam, YM i in. Zmienność międzytaksonomiczna wzorców gęstości kości i zróżnicowana reprezentacja elementów Bovid, Vervid i Equid w zapisie archeologicznym. Am. Antiq . 64 , 343–362.
-
Villa, P. i Mahieu, E. Wzory złamań długich kości człowieka. J. Hum. Evol. 21 , 27–48 (1991).
-
Runge, F. Inwentaryzacja fitolitów opalowych w glebach Afryki Środkowej – ilości, kształty, klasyfikacja i widma. Rev. Palaeobot. Palyno. 107 , 23–53 (1999).
-
Collura, LV i Neumann, K. Fitolity drewna i kory roślin drzewiastych Afryki Zachodniej. Quat. Int. 43 , 142–159 (2017).
-
Fahmy, AG Różnorodność płatowych fitolitów w liściach traw z regionu Sahelu, zachodnia tropikalna Afryka: Tribe Paniceae. Plant Syst. Evol. 270 , 1–23 (2008).
-
Mercader, J., Bennett, T., Esselmont, C., Simpson, S. & Walde, D. Fitolity w roślinach zdrewniałych z lasów Miombo w Mozambiku. Ann. Bot. 104 , 91–113 (2009).
-
Mercader, J. i in. Fitolity Poaceae z Niassa Rift w Mozambiku. J. Archaeol. Nauka. 37 , 1953–1967 (2010).
-
Mercader, J., Bennett, T., Esselmont, C., Simpson, S. i Walde, D. Fitolity glebowe z lasów miombo w Mozambiku. Kwat. Rozdzielczość 75 , 138–150 (2011).
-
Albert, RM, Bamford, MK i Cabanes, D. Tafonomia fitolitów i makroroślin w różnych glebach z wąwozu Olduvai (Tanzania) i jej zastosowanie w próbkach paleoantropologicznych z pliocenu i plejstocenu. Quat. Int. 148 , 78–94 (2006).
-
Madella, M., Alexandre, A. i Ball, T. Międzynarodowy kod nomenklatury fitolitów 1.0. Ann. Bot. 96 , 253–260 (2005).
-
Bennett, KD i Willis, KJ w książce Tracking Environmental Change using Lake Sediments : Terrestrial, Algal, and Siliceous Indicators Vol. 3 (red. Smol, JP, Birks, HJB i Last, WM) (Springer, 2001). Rozdział 2.
-
Levin, NE, Cerling, TE, Passey, BH, Harris, JM i Ehleringer, JR Indeks suchości izotopów trwałych dla środowisk lądowych. Proc. Natl Acad. Sci. 103 , 11201–11205 (2006).
-
Blumenthal, SA i in. Szereg czasowy stabilnych izotopów w zębach ssaków: in situ δ 18 O z najgłębszej warstwy szkliwa. Geochim. Cosmochim. Acta 124 , 223–236 (2014).
-
Roberts, P. i in. Stabilne izotopy kopalnych roślinożerców ujawniają paleośrodowisko homininów w środkowym plejstocenie w „Zielonej Arabii”. Nat. Ecol. Evol. 2 , 1871–1878 (2018).
-
Uno, KT i in. Dieta dużych ssaków i paleoekologia w okresie przejściowym oldowańsko-achulskim w wąwozie Olduvai w Tanzanii na podstawie analizy stabilnych izotopów i zużycia zębów. J. Hum. Evol. 120 , 76–91 (2018).
-
McHenry, LJ Skład fenokryształów jako narzędzie korelacji świeżego i zmienionego tephry, Bed I, Olduvai Gorge, Tanzania. Stratigraphy 2 , 101–115 (2005).
Autorzy:
Komunikacja przyrodnicza tom 12 , Numer artykułu: 3 ( 2021 )
Podziękowanie
Po pierwsze, autorzy doceniają istotny wkład w wiedzę przedstawioną w niniejszym dokumencie społeczności Masajów w wąwozie Oldupai. Praca ta została wsparta przez Canadian Social Sciences and Humanities Research Council w ramach jej Partnership Grant Program nr 895-2016-1017. Tanzania Commission for Science and Technology zatwierdziła tę pracę na mocy pozwolenia nr 2018-112-NA-2018-36. Tanzaniańskie Ministerstwo Zasobów Naturalnych i Turystyki, za pośrednictwem swojego Wydziału Zabytków, udzieliło nam pozwolenia na wykonanie tej pracy (14/2017/2018), a władze w Ngorongoro Conservation Area zezwoliły nam na wejście na teren chroniony (BE.504/620/01/53). Licencję eksportową na materiały przedstawione w tym badaniu uzyskano od Antiquities Division (EA.150/297/01: 5/2018/2019) i tanzańskiego sekretarza wykonawczego Mining Commission (00001258). Pam Akuku i Palmira Saladié są wspierane przez AGAUR (projekt nr 2017 SGR-1040) i URV (2018PFR-URV-B2-91). Program doktorancki Pam Akuku jest finansowany przez Canadian Social Sciences and Humanities Research Council. Nina Jablonsky, Bernard Wood i Michael Lague pomogli nam w identyfikacji szczątków cercopithecine.
Finansowanie
Finansowanie w ramach Open Access umożliwione i zorganizowane przez Projekt DEAL.
Deklaracje etyczne
Konflikty interesów
Autorzy deklarują brak konfliktu interesów.
Informacje dodatkowe
Informacje o recenzjach eksperckich Nature Communications dziękuje anonimowym recenzentom za ich wkład w recenzję ekspercką tej pracy. Raporty recenzentów są dostępne.
Uwaga wydawcy: Springer Nature zachowuje neutralność w kwestii roszczeń jurysdykcyjnych na opublikowanych mapach i powiązań instytucjonalnych.
Informacje uzupełniające
Prawa i uprawnienia
Otwarty dostęp Niniejszy artykuł jest licencjonowany na podstawie licencji Creative Commons Uznanie autorstwa 4.0 Międzynarodowe, która zezwala na używanie, udostępnianie, adaptację, dystrybucję i reprodukcję w dowolnym medium lub formacie, pod warunkiem podania odpowiedniego uznania dla oryginalnych autorów i źródła, podania linku do licencji Creative Commons i wskazania, czy wprowadzono zmiany. Obrazy lub inne materiały stron trzecich w tym artykule są objęte licencją Creative Commons artykułu, chyba że wskazano inaczej w linii kredytowej do materiału. Jeśli materiał nie jest objęty licencją Creative Commons artykułu, a zamierzone użycie nie jest dozwolone przez przepisy ustawowe lub przekracza dozwolone użycie, należy uzyskać zgodę bezpośrednio od właściciela praw autorskich. Aby wyświetlić kopię tej licencji, odwiedź stronę http://creativecommons.org/licenses/by/4.0/ .
O tym artykule

Cytuj ten artykuł
Mercader, J., Akuku, P., Boivin, N. i in. Najwcześniejsze homininy Olduvai eksploatowały niestabilne środowiska ~ 2 miliony lat temu. Nat Commun 12 , 3 (2021). https://doi.org/10.1038/s41467-020-20176-2
Powyższy artykuł był pierwotnie publikowany na stronie Nature Comunications.
Nature Communications to czasopismo o otwartym dostępie, które publikuje wysokiej jakości badania ze wszystkich dziedzin nauk przyrodniczych. Artykuły publikowane w czasopiśmie stanowią ważne postępy o znaczeniu dla specjalistów w każdej dziedzinie.
Współczynnik wpływu czasopisma Nature Communications w okresie 2 lat wynosi 14,7 (2023), liczba pobrań artykułu: 114 944 868 (2023), a od przesłania do pierwszej decyzji redakcyjnej upływa 8 dni (mediana).




 / Piotr Kotlarz")

{kind=link}