")
Prawie 150 lat po pierwszej identyfikacji materiału szkieletowego neandertalczyka zdolności poznawcze i symboliczne tych populacji pozostają przedmiotem intensywnej debaty. Przedstawiamy 99 nowych szczątków neandertalczyka z jaskini Troisième w Goyet (Belgia) datowanych na 40 500–45 500 kal. lat temu. Szczątki zidentyfikowano w drodze multidyscyplinarnego badania, które łączy w sobie morfometrię, tafonomię, stabilne izotopy, datowanie radiowęglowe i analizy genetyczne. Kości neandertalczyka Goyeta wykazują wyraźne modyfikacje antropogeniczne, co stanowi wyraźny dowód na działalność rzeźniczą, a także cztery kości użyte do retuszowania narzędzi kamiennych. Oprócz tego, że Goyet jest pierwszym miejscem, w którym znaleziono wiele kości neandertalczyka używanych do retuszu, nie tylko dostarcza pierwszych jednoznacznych dowodów na kanibalizm neandertalczyka w Europie Północnej, ale także podkreśla znaczną różnorodność zachowań kostnicowych wśród populacji późnego neandertalczyka zamieszkującej ten region w tym okresie bezpośrednio poprzedzających ich zniknięcie.
Podobne treści oglądane przez innych


Wstęp
Neandertalskie praktyki pogrzebowe pozostają w czołówce badań paleoantropologicznych, wywołując gorące debaty po rewizji starych danych i nowych wykopaliskach w kluczowych miejscach, takich jak La Chapelle-aux-Saints 1 , 2 , Roc de Marsal 3 , Saint-Césaire 4 i La Ferrassie 5 . Mówiąc bardziej ogólnie, uwaga badaczy skupiła się na zmienności praktyk pogrzebowych neandertalczyków, aby ocenić ich implikacje poznawcze i symboliczne, zwłaszcza że mogą one zapewnić wgląd w systemy społeczne tej kopalnej grupy ludzkiej 6. Wiadomo, że neandertalczycy chowali swoich zmarłych i kojarzeni są z zachowaniami pogrzebowymi, które często trudno zinterpretować w kontekście paleolitu. Pouczającym przykładem w tym sensie jest Krapina (Chorwacja). Dowody kanibalizmu po raz pierwszy zaproponowano dla tego miejsca już w 1901 roku7 na podstawie fragmentacji i śladów spalenia dużej kolekcji wczesnych szczątków neandertalczyka. Od tego czasu dowody te zostały zakwestionowane przez zwolenników alternatywnych wyjaśnień modyfikacji ludzkich kości, którzy opowiadają się za procesami naturalnymi, podczas gdy inni utrzymują, że manipulacje antropogeniczne najlepiej interpretować w kontekście pochówków wtórnych8. W kilku badaniach poświęconych kanibalizmowi zaproponowano, aby bezpieczna identyfikacja modyfikacji antropogenicznych związanych z tą praktyką uwzględniała dowody na podobne traktowanie szczątków fauny i ludzi w celu wydobycia składników odżywczych 9 , 10 , 11. Oprócz Gran Doliny (poziom TD6; wczesny plejstocen) w Hiszpanii, która dostarczyła najwcześniejszych niekwestionowanych dowodów na kanibalizm 12 , udokumentowano także dalsze przykłady na kilku stanowiskach neandertalczyków w Europie Zachodniej, w tym w El Sidrón i Zafarraya 13 , 14 w Hiszpanii, oraz Moula-Guercy i Les Pradelles 15 , 16 we Francji.
Tutaj dostarczamy nowych danych na temat różnorodności zachowań pogrzebowych neandertalczyków, koncentrując się na niewielkim obszarze ich znanego zasięgu, Europie Północnej, podczas Marine Isotope Stage (MIS) 3 (około 60-30 tysięcy lat temu), w celu zidentyfikowania procesów na małą skalę w tym krótkim okresie, który był świadkiem zniknięcia neandertalczyków17. Przedstawiamy 99 nowych szczątków neandertalczyków zidentyfikowanych niedawno wśród zbiorów z jaskini Troisième w Goyet (Belgia), z których niektóre wykazują modyfikacje antropogeniczne, i omawiamy ich implikacje.
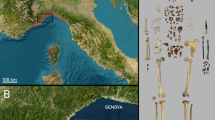
Troisième caverne (lub „Trzecia jaskinia”) w Goyet, wykopana w drugiej połowie XIX i na początku XX wieku oraz ponownie pod koniec lat 90-tych18, jest częścią dużego systemu jaskiń zlokalizowanego w basenie Mosan (rys. uzupełniający S1). Najbardziej rozległe wykopaliska zostały przeprowadzone przez Edouarda Duponta w 1868 roku, który opisał pięć „poziomów faunistycznych” (FBL; ref. 19; uwaga uzupełniająca S1). Jaskinia Troisième przyniosła bogatą sekwencję archeologiczną z osadami z okresu środkowego i górnego paleolitu, zawierającymi artefakty z okresu mousterianu, lincombian-ranisian-jerzmanowician (LRJ), aurignacianu, gravettianu i magdaleńskiego, a także neolitu i okresu historycznego20,21,22,23. To, czy materiał mousterski pochodzi z jednej czy wielu faz okupacji, jest obecnie niemożliwe do rozróżnienia (Nota uzupełniająca S2). Niestety, metody wykopaliskowe nie spełniały dzisiejszych standardów i wydaje się, że poziomy opisane przez Duponta w rzeczywistości reprezentują mieszankę materiału z różnych okresów (np. ref. 24).
Kilka szczątków ludzkich z różnych poziomów zostało opublikowanych przez Dupont19 i Hamy25, chociaż tylko kilka figuruje w Katalogu kopalnych hominidów26, z których wszystkie zostały przypisane do magdaleńskiego. W 2004 r. zidentyfikowaliśmy zarówno fragment żuchwy neandertalczyka, jak i izolowany ząb wśród ludzkiego materiału odzyskanego przez Duponta z jaskini Troisième i obecnie przechowywanego w Królewskim Belgijskim Instytucie Nauk Przyrodniczych (RBINS)27, co czyni Goyet jednym z niewielu północnoeuropejskich miejsc na północ od 50 ° N, które przyniosły szczątki neandertalczyka MIS 3 (Uwaga uzupełniająca S1 i Rys. uzupełniający S1).
Wyniki
Identyfikacja nowych szczątków neandertalczyka w Goyet i ich charakterystyka biogeochemiczna
Ponowna analiza materiału z Goyet obejmowała (I) rewizję ludzkiego materiału szkieletowego, (II) systematyczne sortowanie zbiorów fauny w celu sprawdzenia niezidentyfikowanych szczątków ludzkich (ryc. S2) oraz (III) multidyscyplinarne badanie szczątków ludzkich i ich kontekstu. Zidentyfikowano dwieście osiemdziesiąt trzy szczątki ludzkie z różnych okresów, w tym 96 okazów kości i trzy pojedyncze zęby, które można zidentyfikować jako neandertalskie (Tabela uzupełniająca S1 i Uwagi uzupełniające S3, S4 i S5). Znaczna liczba (n = 47) okazów kości została ponownie dopasowana, zmniejszając całkowitą liczbę izolowanych szczątków neandertalskich do 64 (ryc. 1 i tabela uzupełniająca S2), z których 10 zostało bezpośrednio datowanych radiowęglowo (14C), 15 zostało pobranych do analiz stabilnych izotopów, a 10 do ekstrakcji DNA (tabela 1 i tabela uzupełniająca S3). W oparciu o ich morfologię i cechy morfometryczne, etap rozwoju i stronę dla sparowanych elementów, a także udane odzyskanie sekwencji endogennego mitochondrialnego DNA (mtDNA), minimalną liczbę osobników (MNI) reprezentowanych przez próbkę Goyet szacuje się na pięć (czterech nastolatków/dorosłych i jedno dziecko reprezentowane przez pojedynczy ząb; Uwaga uzupełniająca S5 i rys. uzupełniający S3). Chociaż próbka neandertalczyka obejmuje elementy czaszki i postczaszki (ryc. 1), przy czym najlepiej reprezentowane są kości długie, a kończyny są w większości nieobecne, minimalna liczba elementów (MNE = 35) wskazuje na bardzo niską ogólną reprezentację szkieletu. Najlepiej reprezentowane elementy to, w kolejności malejącej, kość piszczelowa (sześć z ośmiu kości piszczelowych oczekiwanych dla czterech nastolatków/dorosłych, 75% reprezentacji), kość udowa i czaszka (50%), kość ramienna i żuchwa (25%; Tabela uzupełniająca S4).

*Oznacza okazy, które zostały bezpośrednio datowane. Skala = 3 cm.
Chemiczne analizy pierwiastkowe przeprowadzone wraz z analizami stabilnych izotopów wykorzystano do oceny zachowania kolagenu podczas przygotowywania datowania metodą 14C (patrz Metody). Ekologię neandertalczyków Goyeta badano także, wykorzystując skład izotopowy kolagenu kostnego δ 13 C i δ 15 N 28 . Bezpośrednie daty 14 C uzyskane z nowo zidentyfikowanego materiału szkieletowego wskazują, że neandertalczycy z Goyet żyli w okresie ok. 40,5–45,5 kcalBP. Jednakże, gdy wykluczymy najmłodszy wiek, który prawdopodobnie odzwierciedla niewykryte zanieczyszczenie kolagenem kości ( uwaga dodatkowa S6 ), nie możemy wykluczyć możliwości, że neandertalczycy Goyet reprezentują pojedynczą grupę chronologiczną datowaną na ok. 44–45,5 kcalBP. Chociaż wydaje się to najbardziej oszczędną hipotezą, gdy weźmie się pod uwagę indywidualne powiązania kostne, aspekty tafonomiczne i podobne modyfikacje antropogeniczne obserwowane w całej próbie, zachowujemy konserwatywny zakres ok. 40,5–45,5 k k calBP dla neandertalczyków Goyeta w przypadku braku ostatecznych dowodów.
Spośród 10 próbek poddanych analizie genetycznej siedem wykazuje trzy odrębne, kompletne lub prawie kompletne linie mtDNA (oznaczone 1–3 w Tabeli 1 ). Nowo zrekonstruowane mtDNA pochodzące z Goyeta porównano z mtDNA 54 współczesnych ludzi, ośmiu wcześniej zsekwencjonowanych neandertalczyków i jednego osobnika denisowiańskiego 29 , 30 , 31 , 32 , 33 , 34. Powiązania filogenetyczne oceniono przy użyciu drzew maksymalnego oszczędzania i maksymalnego prawdopodobieństwa ( ryc. 2 i ryc. Uzupełniająca S4 ), potwierdzając, że analizowane okazy mieszczą się w znanej różnorodności mtDNA neandertalczyka. MtDNA neandertalczyka Goyeta wydają się najściślej spokrewnione z mtDNA późnego neandertalczyka z Europy Środkowej i Zachodniej, takie jak te ze stanowiska typu neandertalskiego (Niemcy), El Sidrón (Hiszpania) i Vindija (Chorwacja), z których wszystkie wykazują jedynie niewielką zmienność genetyczną pomimo duże odległości geograficzne w porównaniu do współczesnego człowieka. Jak wcześniej sugerowano 31 , może to odzwierciedlać niską efektywną wielkość populacji neandertalczyków w ogóle, a w szczególności późnych neandertalczyków.

Liczby w głównych węzłach gałęzi reprezentują wartości bootstrap po 1000 iteracjach.
Analiza tafonomiczna materiału neandertalskiego Goyeta i modyfikacje antropogeniczne
Ogólnie rzecz biorąc, szczątki neandertalczyka są bardzo fragmentaryczne. Czterdzieści dziewięć procent próbek kości (47 z 96) zostało ponownie dopasowanych do co najmniej jednej innej, przy liczbie próbek na zestaw uzupełniający w zakresie od 2 do 8 (kość piszczelowa I; rys. uzupełniająca S5 ). Zidentyfikowano także kilka przykładów remontów na poziomach od 1 do 3. Żadna z kości neandertalczyka nie jest kompletna, chociaż proksymalny koniec paliczka dłoni (2878–37) jest jedynie nieznacznie nadwyrężony ( ryc. 1). Powierzchnie korowe są dobrze zachowane i wykazują ograniczone modyfikacje postdepozycyjne. Większość złamań kości długich obejmuje zielone przerwy, na co wskazują gładkie brzegi i złamania spiralne 35 . Ślady łuszczenia mogą również świadczyć o złamaniu świeżej kości fragmentu czaszki i kilku żeber (ref. 11 ; ryc. Uzupełniająca S6 ). Chociaż niedźwiedzie mogą pozostawiać takie ślady 36 , obecność śladów nacięć na kilku żebrach (patrz poniżej) sugeruje, że najbardziej oszczędną hipotezą jest to, że są one antropogeniczne. Podejrzewa się również ślady ludzkiego żucia 37 , 38 na paliczkach neandertalczyka, ale są one niejednoznaczne ( rysunek uzupełniający S6 ). Liczne jednoznaczne ślady antropogeniczne na szczątkach neandertalczyka Goyeta można przypisać trzem kategoriom modyfikacji powierzchni kości ( ryc. 3 , 4 , 5 , tabela 2 oraz ryc. uzupełniające S7 i S8 ):
-
Ślady cięcia. Prawie jedna trzecia okazów neandertalczyka nosi ślady nacięć. Lokalizacje nielicznych śladów nacięć zaobserwowanych na kończynie górnej mogą wskazywać na rozczłonkowanie, natomiast te na kończynie dolnej wskazują na migotanie. Kilka śladów nacięć na wewnętrznej i zewnętrznej powierzchni żeber może być związanych z wypatroszeniem, rozczłonkowaniem klatki piersiowej i usunięciem mięśni klatki piersiowej. Dodatkowe nacięcie po środkowej stronie żuchwy, w pobliżu kłykcia żuchwy, wydaje się wskazywać na rozczłonkowanie.
-
Wyróżniono dwa typy śladów uderzeń (nacięcia i wgłębienia). Zaobserwowane tylko w jednym promieniu wzdłuż kilku kości udowych i piszczelowych, wcięcia są prawdopodobnie związane ze złamaniem świeżych trzonów kości i ekstrakcją szpiku. Wgłębienia po uderzeniu są częste i prawdopodobnie wskazują na nieudane próby złamania kości. Na ośmiu kościach (np. kości udowej I, ryc. 5 ) zidentyfikowano także nacięcia i wgłębienia opukowe .
-
Retusz śladów. Ślady te, znalezione na kości udowej i trzech kościach piszczelowych ( ryc. uzupełniające S9 – S12 ), wynikają z retuszowania krawędzi narzędzi kamiennych. Fakt, że żaden z dotkniętych obszarów nie nakłada się na sąsiednie fragmenty, sugeruje, że kości prawdopodobnie najpierw uległy pęknięciu szpiku. Kość udowa III przedstawia dwie strefy retuszu na powierzchni przedniej i tylno-przyśrodkowej, obie zlokalizowane w połowie trzonu. Co ciekawe, ślady znalezione na kościach piszczelowych znajdują się w tych samych obszarach trzonu na wszystkich trzech kościach (powierzchnia tylna lub tylno-przyśrodkowa w połowie trzonu). Retusze są wykonane na czterech różnych kościach neandertalczyka, które reprezentują co najmniej trzy z czterech dorastających / dorosłych osobników neandertalczyków ( uwaga dodatkowa S5 ).

Zobacz dodatkową ryc. S8 dla poszczególnych kości neandertalczyka z modyfikacjami antropogenicznymi. Diagramy szkieletu zmodyfikowano z https://en.wikipedia.org/wiki/File:Human_skeleton_front_en.svg i https://en.wikipedia.org/wiki/File:Human_skeleton_back_en.svg przy użyciu programu Adobe Illustrator CS4 v. 14.0.0.

( a ) kość udowa III w widoku z przodu; ( b1 , c1 ) zdjęcia z bliska; ( b2 , c2 ) obrazy uzyskane przy użyciu minidomy (patrz Metody).

( a ) kość udowa I w widoku od tyłu; ( b1 , c1 ) zdjęcia z bliska; ( b2 , c2 ) obrazy uzyskane przy użyciu minidomy (patrz Metody).
Chociaż retusze kości zwierzęcych były powszechne w europejskim kontekście środkowego paleolitu (np. ref. 39 , 40 , 41 ), Goyet jest jednym z zaledwie czterech stanowisk (Krapina w Chorwacji 42 , La Quina i Les Pradelles we Francji 43 , 16 ), w których uzyskano retusze na neandertalskich elementach szkieletu i podeszwie dały wiele przykładów ( Tabela 3 ). W Krapinie i Les Pradelles jako retusze wykorzystano fragmenty trzonu kości udowej, podczas gdy w La Quina wykorzystano fragment ciemieniowy. Według kryteriów zaproponowanych przez Mallye i in . 40 , półfabrykaty użyte do retuszerów Goyeta wykonanych na kościach neandertalczyka były najprawdopodobniej zielone ze względu na brak obszarów złuszczonych, a ponadto na dwóch z pięciu obszarów retuszerów widoczne są skoncentrowane i nałożone na siebie ślady, co sugeruje długotrwałe użytkowanie. Prostoliniowa morfologia znaków przemawia również za wykorzystaniem kości do retuszowania odłupków krzemiennych, najpowszechniejszego surowca spotykanego w Goyet.
Porównawcza analiza tafonomiczna fauny jaskini Troisième
Ze względu na duży rozmiar kolekcji fauny Goyeta (> 30 000 okazów) zbadano tylko próbkę z wykopalisk Duponta (patrz metody; rys. uzupełniająca S2 i tabela uzupełniająca S5 ). Analizowany materiał szkieletowy to głównie fragmenty trzonów kości długich różnych gatunków, które zostały zmieszane w obrębie kolekcji i nie sprawiały wrażenia wcześniej sortowanych. Skupiliśmy się na szczątkach z poziomów 3 i 2, które dostarczyły szczątki neandertalczyka, oraz na materiale z tych samych tac do przechowywania, w których znajdowały się szczątki ludzkie, aby uzyskać przegląd powiązanego spektrum fauny i ocenić strategie pozyskiwania żywności i zarządzania nią. Zdecydowanie najczęstszym gatunkiem w badanym zbiorowisku są konie i renifery (86% z 1556 zidentyfikowanych okazów; tabela uzupełniająca S5 ). Nie zaobserwowano śladów zębów gryzoni, szczątki mięsożerców są stosunkowo rzadkie, a uszkodzenia mięsożerców są niezwykle rzadkie w przypadku szczątków neandertalczyka, koni i reniferów ( Tabela 2 ), co wskazuje, że mięsożercy mieli ograniczony dostęp do materiału kostnego.
Profile anatomiczne ujawniają liczne podobieństwa między próbką neandertalczyka z jednej strony a koniem i reniferem z drugiej ( tabela uzupełniająca S6 i ryc. Uzupełniająca S13 ). Ze wszystkich trzech gatunków najliczniejszym elementem jest kość piszczelowa, natomiast szkielet osiowy oraz kończyny kończyn przednich i tylnych są słabo reprezentowane. Kości kończyny tylnej są lepiej reprezentowane u wszystkich trzech gatunków w porównaniu z elementami kończyny przedniej, dotyczy to szczególnie materiału neandertalskiego. Jedyną zauważalną różnicą między szczątkami fauny i neandertalczyka jest duża reprezentacja elementów czaszkowych w przypadku tego ostatniego. Niestety brak danych kontekstowych uniemożliwia analizę przestrzennego rozmieszczenia zarówno fauny, jak i szczątków neandertalczyka w jaskini Troisième.
Najbardziej intensywnie przetworzonymi elementami neandertalskimi są kości udowe i piszczelowe ( rysunek uzupełniający S7 ), które są również kośćmi o najwyższej zawartości składników odżywczych (mięso i szpik). Ten sam wzór udokumentowano w przypadku kości koni i reniferów. Ogólnie rzecz biorąc, ślady antropogeniczne na szczątkach neandertalczyka odpowiadają tym najczęściej odnotowanym na materiale fauny ( rysunki uzupełniające S14 – S16 ). Wszystkie trzy taksony były intensywnie eksploatowane, wykazując ślady skórowania, filetowania, dezartykulacji i ekstrakcji szpiku. Jednakże szczątki neandertalczyka wyróżniają się dużą liczbą jamek udarowych ( tabela 2 ), co można powiązać z grubą strukturą korową długich kości neandertalczyka. Chociaż na szczątkach neandertalczyka nie widać śladów spalenia, nie można wykluczyć, że mogły zostać upieczone lub ugotowane. Duża liczba śladów nacięć i fakt, że można było z powodzeniem wyodrębnić DNA, stoją jednak w sprzeczności z tą możliwością 44 , 45 , 46 . Wreszcie, podobnie jak odnotowano w innych miejscach 40 , 41 , 47 , retusze neandertalskie wykonuje się na fragmentach gęstych kości o właściwościach mechanicznych porównywalnych do kości konia i renifera. W Goyet, podobnie jak na kilku stanowiskach ze środkowego paleolitu we Francji, wybrano duże fragmenty kości zwierząt średnich i dużych 40 , 41 , 48 , 49 , 50 , 51 . Spośród materiału Goyeta neandertalskiego wyselekcjonowano także fragmenty największe i najgrubsze, jak miało to miejsce w Les Pradelles 16 i Krapina 42 . Co ciekawe, wśród półproduktów retuszerskich wybranych przez neandertalczyków w Scladina znajdowała się także kość udowa i piszczele niedźwiedzi jaskiniowych 52 .
Zaobserwowane wzorce eksploatacji fauny można interpretować jako selektywny transport na stanowisko składników bogatych w mięso i szpik kostny, które następnie podlegały intensywnej obróbce. Jednakże ten widoczny wzór może odzwierciedlać tendencję do faworyzowania największych i najłatwiejszych do zidentyfikowania fragmentów. Podobieństwa w śladach antropogenicznych zaobserwowane na kościach neandertalczyka, koni i reniferów sugerują jednak podobne wzorce przetwarzania i konsumpcji w przypadku wszystkich trzech gatunków.
Dyskusja
Nasze wyniki pokazują, że neandertalczycy z jaskini Troisième w Goyet zostali zarżnięci, a hipoteza ich wykorzystania jako źródła pożywienia jest najbardziej prawdopodobnym wyjaśnieniem obserwowanych modyfikacji powierzchni kości. Goyet dostarcza pierwszych jednoznacznych dowodów na kanibalizm neandertalczyków w Europie Północnej, a biorąc pod uwagę daty uzyskane na szczątkach neandertalczyków, najprawdopodobniej zostały one przetworzone przez ich pobratymców neandertalczyków, ponieważ nie wiadomo, aby współcześni ludzie byli w tym czasie w regionie17,23. Jednak dostępne dane uniemożliwiają ustalenie, czy modyfikacje zaobserwowane na neandertalskim materiale szkieletowym reprezentują praktyki symboliczne, czy po prostu wynikają z przetwarzania bezpośrednio dostępnych źródeł żywności. Ponadto, Goyet jest pierwszym stanowiskiem, które dostarczyło wielu neandertalskich retuszerów kości. Zaproponowano, że półfabrykaty retuszerów ze środkowego paleolitu były produktami ubocznymi przetwarzania tusz do spożycia40,41, które mogły zostać wybrane do ponownego użycia51. Dostępne dane nie pozwalają nam zaproponować innego scenariusza dla retuszerów Goyet wykonanych na kościach neandertalczyków. Jednak świeżość użytych półfabrykatów sugeruje, że neandertalczycy mogli być świadomi, że używają ludzkich szczątków. Nie można poświadczyć, czy była to część działalności symbolicznej, czy też wywołana motywacją funkcjonalną, jak miało to miejsce w przypadku neandertalskiego retuszera z La Quina43.
Chociaż późni neandertalczycy z Goyet datowani są na 40,5-45,5 ky calBP, brak wiarygodnych informacji kontekstowych uniemożliwia powiązanie ich z którymkolwiek z technokompleksów z tego stanowiska. Jednak zespoły mousterskie są znane ze stanowisk w basenie Mosan, jak w jednostce 1A Scladina53, położonej zaledwie 5 km od Goyet, warstwie CI-8 jaskini Walou54 i warstwie II Trou de l’Abîme w Couvin55 (Nota uzupełniająca S2). Podczas gdy LRJ jest znany z dwóch miejsc w Belgii, Spy i Goyet, a jego pierwsze pojawienie się w innych miejscach datowane jest na około 43-44 ky calBP23,56), obecnie nie są dostępne żadne wiarygodne informacje na temat jego regionalnej chronologii. Biorąc pod uwagę bezpośrednie daty 14C uzyskane dla neandertalczyków z Goyet, niemożliwe jest bezpieczne powiązanie ich z okupacją (okupacjami) Mousterian lub LRJ.
Jeśli chodzi o późne neandertalskie praktyki pogrzebowe w regionie, cztery stanowiska w promieniu około 250 km wokół Goyet wyprodukowały neandertalskie szczątki wiarygodnie datowane na 50-40 ky calBP (rys. uzupełniający S1). Co ciekawe, żadne z tych stanowisk nie dostarczyło dowodów na traktowanie zwłok podobnych do tych udokumentowanych dla Goyet. Dwa belgijskie stanowiska, Walou Cave i Trou de l’Abîme, wyprodukowały odpowiednio przedtrzonowiec i trzonowiec55,57. Chociaż niemożliwe jest wywnioskowanie sygnatury behawioralnej reprezentowanej przez te szczątki, biorąc pod uwagę ich stan zachowania, jest bardzo mało prawdopodobne, że obejmowały one praktyki pogrzebowe, w tym pochówek. W Niemczech neandertalczycy z Feldhofer, w tym neandertalczyk 1, są prawdopodobnie powiązani z „grupą Keilmesser”, późnopaleolitycznym technokompleksem58,59 nieznanym w Goyet (uwaga uzupełniająca S2). Neandertalczyk 1 zawiera elementy szkieletu czaszki i postczaszki pojedynczego osobnika. Pomimo śladów cięcia na czaszce, obojczyku i łopatce, kości długie są nienaruszone, a uszkodzenie wciąż połączonych elementów szkieletu podczas ich odzyskiwania wskazuje, że przynajmniej część szkieletu mogła być pierwotnie połączona anatomicznie60,61. Wreszcie, w Spy, bezpośrednie daty uzyskane na dwóch dorosłych neandertalczykach umieszczają je w obecnej chronologii LRJ62, chociaż związek między ludzkimi szczątkami a tym technokompleksem jest niepewny z powodu braku informacji kontekstowych. Niedawna ponowna ocena okazów Spy i ich kontekstu sugeruje, że obie osoby zostały pochowane63. Warto też zauważyć, że najbardziej kompletny osobnik, Spy II, został pierwotnie opisany jako kompletny szkielet znaleziony w pozycji skurczonej. Co więcej, kompletność szkieletu i brak zmian po złożeniu sugerują, że ciało zostało szybko zabezpieczone63.
W zachowaniach pogrzebowych populacji późnego neandertalu z Europy Północnej jest widoczna znaczna różnorodność, prawdopodobnie obejmująca zarówno pierwotne, jak i wtórne depozyty, wraz z innymi rodzajami praktyk, w tym kanibalizmem. Pomimo niskiej różnorodności genetycznej wśród populacji późnego neandertalu, obecność różnych technokompleksów z późnego środkowego paleolitu, a także LRJ, sugeruje jednak znaczną zmienność behawioralną wśród tych grup w Europie Północnej.
Metody
Ocena kolekcji
Do oceny zbiorów Goyeta uwzględniono materiały znajdujące się w RBINS i Królewskich Muzeach Sztuki i Historii (RMAH) w Brukseli, które pochodzą z jaskini Troisième, a także zbiory z Grand Curtius Museum (Liège), Cercle d’Histoire et d’Archéologie du Pays de Genappe (Genappe) i Préhistosite de Ramioul (Ramioul), którego pochodzenie jest mniej pewne. Prezentowane tu szczątki neandertalczyka odnaleziono jedynie w dwóch pierwszych kolekcjach. System numeracji okazów i ich pochodzenie omówiono w nocie dodatkowej S4.
Badanie tafonomiczne
Po określeniu składu zespołu fauny pobranego z kolekcji Duponta ( tabela dodatkowa S5 ) za pomocą mikroskopu jednoocznego (× 10) zaobserwowano łącznie 442 szczątki koni i 287 reniferów, podobnie jak wszystkie szczątki neandertalczyka Goyeta. Modyfikacje tafonomiczne i antropogeniczne zarejestrowano i narysowano na mapach anatomicznych ( rysunki uzupełniające S8 i S14 – S16). Rozróżniano ślady nacięć i podeptań ze względu na ich morfologię i umiejscowienie na kościach 64 . Jako ślady udarowe zarejestrowano jedynie jednoznaczne nacięcia z ujemną blizną płatkową 65 , 66 wykonaną na świeżej kości 35 oraz wgłębienia udarowe (pozostawione po uderzeniach po ref. 66 ). Wszystkie zidentyfikowane retuszery kości to fragmenty trzonów kości długich, które wykazują ślady opisane przez Mallye i in . 40 . Na koniec zarejestrowano ślady zębów, stosując typologię Binforda 67 . Zaobserwowano jedynie doły i nacięcia. Niektóre z tych pestek mogły powstać w wyniku przeżuwania przez człowieka 38 , 67 , 68 , 69 , ale nie są one na tyle charakterystyczne, aby zdecydowanie odróżnić je od śladów pozostawionych przez mięsożerców. Za Bello i in . 70 , antropogeniczne modyfikacje zarejestrowane na szczątkach neandertalczyka udokumentowano za pomocą rysunków, zdjęć z bliska i obrazowania w wysokiej rozdzielczości. Obrazy o wysokiej rozdzielczości ( ryc. 4 i 5 oraz ryc. uzupełniające S8 – S12 ) uzyskano przy użyciu minidome, cyfrowego urządzenia do obrazowania opracowanego przez VISICS w KULeuven ( http://www.minidome.be ). Kopuła oparta na technice wielomianowego mapowania tekstury składa się z 260 diod LED i jednej stałej kamery, która rejestruje obraz z każdą diodą LED zapaloną indywidualnie. Wyniki pozwalają na interaktywne wyświetlenie obiektu przy różnym oświetleniu, aby odsłonić wszystkie szczegóły jego powierzchni. Dodatkowo modele 3D retuszerów wykonane na kościach neandertalczyka uzyskane za pomocą systemu pomiaru 3D światłem białym ( http://www.mechscan.co.uk/ ) są dostępne pod adresem http://virtualcollections.naturalsciences.be/virtual-collections/ antropologia-prehistoria/szczątki-ludzkie/goyet.
Dobór i przygotowanie próbek do analiz izotopowych i genetycznych
Wszystkie próbki, z których pobrano próbki, były nieobrobione (sklejone lub lakierowane), nowo zidentyfikowane kości neandertalczyka, z wyjątkiem zęba 2878–2D (patrz tabela 1 ). Próbki przed pobraniem zeskanowano lub zeskanowano i uformowano przy użyciu elastomeru silikonowego DC-3481, a zdjęcia wykonano zarówno przed, jak i po. Kolagen ekstrahowano w Centrum Badań Izotopów Uniwersytetu w Groningen (CIO, Holandia) i grupie roboczej Biogeologii na Wydziale Nauk o Ziemi Uniwersytetu w Tybindze (Niemcy; patrz tabela uzupełniająca S3 ). Datowanie radiowęglowe przeprowadzono w CIO; stabilne izotopy i analizy genetyczne przeprowadzono na Uniwersytecie w Tybindze.
Analizy izotopowe
Ekstrakcja kolagenu w CIO przebiegała zgodnie z procedurą opracowaną przez Longin 71 , z dodatkową obróbką chemiczną przy zastosowaniu standardowych procedur 72 . Ekstrakcję kolagenu na Uniwersytecie w Tybindze przeprowadzono zgodnie z procedurą zmodyfikowaną na podstawie Longin 71 opisaną przez Bocherensa i in . 73 . Do pomiarów stabilnych izotopów ( 13 C, 15 N) wykorzystano analizator elementarny NC 2500 podłączony do spektrometru mas Thermo Quest Delta+XL. Stopień chemicznego utrwalenia kolagenu wyraża się stosunkiem atomowym C coll : N coll , którego dopuszczalny zakres zmienności wynosi 2,9–3,6 74, natomiast zawartość azotu (N coll ) powinna przekraczać 5% 75 . Zawartość węgla w ekstrahowanym kolagenie waha się od 29,5 do 47,1%, a zawartość azotu od 10,1 do 17,0% ( tabela dodatkowa S3 ), przy czym obie wartości mieszczą się w zakresie świeżego kolagenu 76 . Stosunki atomowe C coll : N coll obejmują wąski zakres (3,2–3,4), wskazując wyjątkowo dobrze zachowany kolagen dla wszystkich próbek kości (patrz uwaga dodatkowa S6 w celu omówienia zęba 2878–2D). Następnie kolagen został wysłany do CIO w celu datowania metodą 14 C przez AMS 77 .
Analizy genetyczne
Pobrano próbki z dziesięciu próbek ( Tabela 1 ). DNA ekstrahowano 78 z proszku kostnego i konwertowano do podwójnie indeksowanych bibliotek genetycznych 79 , 80 . DNA mitochondrialne (mtDNA) wzbogacono przy użyciu techniki wychwytywania przynęty 81 i zsekwencjonowano na platformie sekwencjonowania nowej generacji (Illumina HiSeq). Po filtrowaniu jakości i łączeniu odczytów ze sparowanych końców 82 , zmodyfikowaną wersję narzędzia mapującego BWA i pakietu SAMtools w połączeniu z niestandardowym iteracyjnym asemblerem mapującym 30 , 83 , 84 , 85 zastosowano w celu dopasowania odczytów do referencyjnej sekwencji mtDNA neandertalczyka ( dodatkowe Tabela S7 ). Odczyty z trzech próbek o niskim pokryciu zostały również dopasowane do współczesnej ludzkiej sekwencji referencyjnej mtDNA (rCRS), aby wykluczyć błędy referencyjne ( tabela uzupełniająca S8 ). Nowo zrekonstruowane kompletne lub prawie kompletne (tj. co najmniej 98% kompletne) genomy mitochondrialne porównano z 63 innymi sekwencjami mtDNA homininy w drzewach genowych ( ryc. 2 i ryc. Uzupełniająca S4 ) w celu oceny ich umiejscowienia filogenetycznego i powiązań genetycznych między grupami ( uwaga dodatkowa S7 ). Autentyczność uzyskanych sekwencji mitochondrialnych jako endogennego starożytnego DNA zweryfikowano poprzez analizę typowych wzorów uszkodzeń starożytnego DNA (ref. 86 i ryc. uzupełniająca S17 ), a także oszacowanie procentu zanieczyszczenia współczesnym ludzkim DNA (ref. 30 i tabela uzupełniająca S7 ) . Na koniec odfiltrowano uszkodzone cząsteczki DNA wskazujące na starożytne pochodzenie i wykorzystano je do zbudowania nowych sekwencji konsensusowych mtDNA. Zostały one wspólnie przeanalizowane z tymi samymi 63 mtDNA w celu potwierdzenia przypisanego umiejscowienia filogenetycznego ( rysunki uzupełniające S18 i S19 ).
Autorzy: Hélène Rougier, Isabelle Crevecoeur, Cedric Beauval, Cosimo Post, Damien Flas , Christoph Wißing, Anja Furtwängler, Mietje Germonpré, Asier Gómez-Olivenci, Patryk Semal, Johannes van der Plicht, Hervé Bocherens i Johannes Krause.
Dodatkowe informacje
Kody dostępu: Siedem sekwencji mtDNA neandertalczyka opisanych w tym artykule zostało zdeponowanych w GenBank i są dostępne pod numerami dostępu KX198082-KX198088.
Jak cytować ten artykuł : Rougier, H. i in . Kanibalizm neandertalski i kości neandertalczyka używane jako narzędzia w Europie Północnej. Nauka. Rep. 6 , 29005; doi: 10.1038/srep29005 (2016).
Bibliografia
-
Rendu, W. i in. Dowody potwierdzające celowy pochówek neandertalczyka w La Chapelle-aux-Saints. Proc. Natl. Acad. Nauka. USA 111 , 81–86 (2014).
-
Dibble, HL i in. Krytyczne spojrzenie na dowody z La Chapelle-aux-Saints potwierdzające celowy pochówek neandertalczyka. J. Archaeol. Nauka. 53 , 649–657 (2015).
-
Sandgathe, DM, Dibble, HL, Goldberg, P. i McPherron, SP Dziecko neandertalskie Roc de Marsal: ponowna ocena jego statusu jako celowego pochówku. J. Hum. Ewolucja 61 , 243–253 (2011).
-
Bachellerie, F. i in. Wstępne wyniki trwających wykopalisk w La Roche à Pierrot w Saint-Césaire. XVII Światowy Kongres UISPP (Burgos, 1–7 września 2014).
-
Gómez-Olivencia, A., Crevecoeur, I. & Balzeau, A. La Ferrassie 8 Reaktywacja dziecka neandertalskiego: nowe szczątki i ponowna ocena oryginalnej kolekcji. J. Hum. Ewolucja 82 , 107–126 (2015).
-
Pettitt, PB Neandertalczyk zmarły: badanie zmienności grobowców w środkowej Eurazji paleolitu. Przed rolnictwem 2002/1 , 1–26 (2002).
-
Gorjanović-Kramberger, K. Der paläolithische Mensch und seine Zeitgenossen aus dem Diluvium von Krapina in Kroatien I. Mitt. Antropol. Gesella. Wien 31 , 164–197 (1901).
-
Orschiedt, J. Der Fall Krapina – neue Ergebnisse zur Frage von Kannibalismus beim Neandertaler. Quartär 55 , 63–81 (2008).
-
Villa, P. i in. Kanibalizm w neolicie. Nauka 233 , 431–437 (1986).
-
White, TD Prehistoryczny kanibalizm w Mancos 5MTUMR-2346 (Princeton University Press, 1992).
-
Boulestin, B. Approche Taphonomique des Restes Humains. Le Cas des Mésolithiques de la Grotte des Perrats et le Problème du Cannibalisme en Préhistoire Récente Européenne (BAR IS 776, Archaeopress, 1999).
-
Fernández-Jalvo, Y., Díez, JC, Cáceres, I. i Rosell, J. Kanibalizm ludzki we wczesnym plejstocenie Europy (Gran Dolina, Sierra de Atapuerca, Burgos, Hiszpania). J. Hum. Ewolucja 37 , 591–622 (1999).
-
Rosas, A. i in. Paleobiologia i morfologia porównawcza próbki późnego neandertalczyka z El Sidrón w Asturii w Hiszpanii. Proc. Natl. Acad. Nauka. USA 103 , 19266–19271 (2006).
-
Barroso Ruiz, C. i in. W El Pleistoceno Superior de la Cueva del Boquete de Zafarraya. (współpraca Barroso Ruiz, C.) 389–419 (Archeología Monografías, Junta de Andalucía, 2003).
-
Defleur, A., White, T., Valensi, P., Slimak, L. i Crégut-Bonnoure, E. Kanibalizm neandertalczyka w Moula-Guercy, Ardèche, Francja. Nauka 286 , 128–131 (1999).
-
Mussini, C. Les Restes Humains Moustériens des Pradelles (Marillac-le-Franc, Charente, Francja): Etude Morphométrique et Réflexions sur un Aspect Comportemental des Néandertaliens. Te doktoraty (Université Bordeaux 1, 2011).
-
Higham, T. i in. Czas i czasoprzestrzenny wzór zniknięcia neandertalczyka. Natura 512 , 306–309 (2014).
-
Toussaint, M. W neandertalczykach w Europie. (reż. Demarsin, B. & Otte, M.) 115–134 (ERAUL 117, ATVATVCA 2, 2006).
-
Dupont, E. L’Homme wisiorek les Ages de la Pierre dans les Environs de Dinant-sur-Meuse, wydanie 2ème (Muquardt, C. Ed., 1872).
-
Ulrix-Closset, M. Le Paléolithique Moyen dans le Bassin Mosan en Belgique (Universa, 1975).
-
Otte, M. Le Paléolithique supérieur ancien en Belgique. Monogr. Archéol. Nat. 5 , 1–684 (1979).
-
Dewez, M. Le Paléolithique Supérieur Récent dans les Grottes de Belgique (Université Catholique de Louvain, 1987).
-
Flas, D. Przejście od środkowego do górnego paleolitu w Europie Północnej: okres linkombiańsko-ranizyjsko-jerzmanowicki i problematyka akulturacji ostatnich neandertalczyków. Światowy Archeol. 43 , 605–627 (2011).
-
Germonpré, M. Rekonstrukcja przestrzennego rozmieszczenia pozostałości fauny z Goyet w Belgii. Notae Praehistoricae 21 , 57–65 (2001).
-
Hamy, E.-T. Sur quelques ossements humains découverts dans la troisième caverne de Goyet, près Namèche (Belgique). Byk. Towarzystwo Antrop. Paryż 8 , 425–435 (1873).
-
Twiesselmann, F. W katalogu kopalnych hominidów – część 2: Europa. (red. Oakley, KP, Campbell, BG i Molleson, TI) 5–13 (Powiernicy Brytyjskiego Muzeum Historii Naturalnej, 1971).
-
Rougier, H., Crevecoeur, I., Semal, P. i Toussaint, M. In Paléolithique Moyen en Wallonie. Kolekcja La Louis Eloy. (red. Di Modica, K. i Jungels, C.) 173 (Collections du Patrimoine Culturel de la Communauté française 2, 2009).
-
Wißing, C. i in. Izotopowe dowody na ekologię diety późnych neandertalczyków w północno-zachodniej Europie. Kwat. Wewnętrzne 10.1016/j.quaint.2015.09.091 (2015).
-
Ingman, M., Kaessmann, H., Pääbo, S. i Gyllensten, U. Zmienność genomu mitochondrialnego i pochodzenie współczesnych ludzi. Natura 408 , 708–713 (2000).
-
Green, RE i in. Kompletna sekwencja genomu mitochondrialnego neandertalczyka określona metodą wysokowydajnego sekwencjonowania. Komórka 134 , 416–426 (2008).
-
Briggs, AW i in. Ukierunkowane wyszukiwanie i analiza pięciu genomów mtDNA neandertalczyka. Nauka 325 , 318–321 (2009).
-
Krause, J. i in. Kompletny genom mitochondrialnego DNA nieznanego hominina z południowej Syberii. Natura 464 , 894–897 (2010).
-
Prüfer, K. i in. Pełna sekwencja genomu neandertalczyka z gór Ałtaj. Natura 505 , 43–49 (2014).
-
Skoglund, P. i in. Oddzielenie endogennego starożytnego DNA od współczesnego skażenia u syberyjskiego neandertalczyka. Proc. Natl. Acad. Nauka. USA 111 , 2229–2234 (2014).
-
Villa, P. i Mahieu, E. Wzory pękania ludzkich kości długich. J. Hum. Ewolucja 21 , 27–48 (1991).
-
Arilla, M., Rosell, J., Blasco, R., Domínguez-Rodrigo, M. i Pickering, TR Podstawy „niedźwiedzia”: badania aktualne nad Ursus arctos arctos w hiszpańskich Pirenejach i ich implikacje dla paleontologii i archeologii. PLoS ONE 9 , e102457 (2014).
-
Bello, SM, Saladié, P., Cáceres, I., Rodríguez-Hidalgo, A. & Parfitt, SA Rytualny kanibalizm górnego paleolitu w jaskini Gough (Somerset, Wielka Brytania): Szczątki ludzkie od stóp do głów. J. Hum. Ewolucja 82 , 170–189 (2015).
-
White, TD i Toth, N. W tchnięciu życia w skamieniałości: badania tafonomiczne na cześć mózgu CK (Bob). (red. Pickering, TR, Schick, K. i Toth, N.) 281–296 (Stone Age Institute Press, 2007).
-
Patou-Mathis, M. wyd. W Fiches de la Commission de Nomenclature sur l’Industrie de l’Os Préhistorique, X. 11–19 (Société Préhistorique Française, 2002).
-
Mallye, J.-B. i in. Mousterscy retuszerzy kości z jaskini Noisetier: eksperymenty i identyfikacja śladów. J. Archaeol. Nauka. 39 , 1131–1142 (2012).
-
Daujeard, C. i in. Retuszerzy kości ze środkowego paleolitu w południowo-wschodniej Francji: zmienność i funkcjonalność. Kwat. Wewnętrzne 326–327 , 492–518 (2014).
-
Patou-Mathis, M. Analyze taphonomique et paléoethnographique du matériel osseux de Krapina (Chorwacja): nouvelles données sur la faune et les restes humains. Préhistoire Européenne 10 , 63–90 (1997).
-
Verna, C. i d’Errico, F. Najwcześniejsze dowody użycia ludzkiej kości jako narzędzia. J. Hum. Ewolucja 60 , 145–157 (2011).
-
Villa, P. i in. Un cas de kanibalizm au Néolithique. Gallia préhistoire 29 , 143–171 (1986).
-
Abe, Y. Wzorce łowieckie i rzeźnicze Evenków w północnej Transbaikalii w Rosji. Praca doktorska (Stony Brook University, 2005).
-
Thompson, T. Archeologia kremacji: spalone szczątki ludzkie w badaniach pogrzebowych (Oxbow Books, 2015).
-
Van Kolfschoten, T., Parfitt, SA, Serangeli, J. i Bello, SM Narzędzia kostne z dolnego paleolitu z „Horyzontu Włóczni” w Schöningen (Niemcy). J. Hum. Ewolucja 89 , 226–263 (2015).
-
Beauval, C. In Le Site Paléolithique de Chez-Pinaud à Jonzac, Charente-Maritime . (red. Airvaux, J.) 125–156 (Préhistoire du Sud Ouest Suppl. 8, 2004).
-
Jaubert, J. i in. W Les Sociétés du Paléolithique dans un Grand Sud-Ouest de la France: Nouveaux Gisements, Nouveaux Résultats, Nouvelles Méthodes. Journées SPF, Université Bordeaux 1, Talence, 24–25 listopada. 2006 (reż. Jaubert, J., Bordes, J.-G. i Ortega, I.) 203–243 (Mémoire XLVII de la Société Préhistorique Française, 2008).
-
Costamagno, S., Meignen, L., Beauval, C., Vandermeersch, B. & Maureille, B. Les Pradelles (Marillac-le-Franc, Francja): Obóz polowań na renifery mousterskie? J.Anthropol. Archeol. 25 , 466–484 (2006).
-
Rosell, J. i in. Recykling kości w środkowym plejstocenie: Kilka refleksji z Gran Doliny TD10-1 (Hiszpania), Jaskini Bolomor (Hiszpania) i Jaskini Qesem (Izrael). Kwat. Wewnętrzne 361 , 297–312 (2015).
-
Abrams, G., Bello, SM, Di Modica, K., Pirson, S. i Bonjean, D. Kiedy neandertalczycy używali szczątków niedźwiedzia jaskiniowego ( Ursus spelaeus ): Retusze kości z bloku 5 jaskini Scladina (Belgia). Kwat. Wewnętrzne 326–327 , 274–287 (2014).
-
Di Modica, K. Les Productions Lithiques du Paléolithique Moyen en Belgique: Variabilité des Systèmes d’Acquisition et des Technologies en Odpowiedź na Mosaïque d’Environnements Contrastés. Te doktoraty (MNHN, Université de Liège, 2010).
-
Pirson, S. i in. Kontekst chronostratygraficzny przejścia od środkowego do górnego paleolitu: najnowsze dane z Belgii. Kwat. Wewnętrzne 259 , 78–94 (2012).
-
Toussaint, M. i in. Drugi dolny prawy trzonowiec mleczny neandertalczyka z Trou de l’Abîme w Couvin w Belgii. J. Hum. Ewolucja 58 , 56–67 (2010).
-
Cooper, LP i in. Stacja na świeżym powietrzu z wczesnego górnego paleolitu i legowisko hien środkowodewenskich w Grange Farm, Glaston, Rutland, Wielka Brytania. Proc. Prehist. Towarzystwo 78 , 73–93 (2012).
-
Toussaint, M. In La Grotte Walou à Trooz (Belgia). Fouilles de 1996 do 2004 . Głośność 2 . (reż. Draily, C., Pirson, S. & Toussaint, M.) 148–161 (Etudes et Documents – Archéologie 21, 2011).
-
Schmitz, RW i in. Ponowna wizyta w miejscu typu neandertalskiego: interdyscyplinarne badania szczątków szkieletowych z doliny Neandera w Niemczech. Proc. Natl. Acad. Nauka. USA 99 , 13342–13347 (2002).
-
Hillgruber, KF W neandertalczyku 1856–2006. (red. Schmitz, R.) 111–144 (Rheinische Ausgrabungen 58, 2006).
-
Schmitz, RW i Pieper, P. Schnittspuren i Kratzer. Anthropogene Veränderungen am Skelett des Urmenschenfundes aus dem Neandertal – Vorläufige Befundaufnahme. Rheinisches Landesmuseum Bonn 2 , 17–19 (1992).
-
Auffermann, B. & Orschiedt, J. Die Neandertaler, eine Spurensuche (Konrad Theiss Verlag, 2002).
-
Semal, P. i in. Nowe dane na temat późnych neandertalczyków: bezpośrednie datowanie skamieniałości belgijskiego szpiega. Jestem. J.Fiz. Antrop. 138 , 421–428 (2009).
-
Maureille, B., Toussaint, M. i Semal, P. W jaskini szpiegowskiej. 125 lat badań multidyscyplinarnych w Betche aux Rotches (Jemeppe-sur-Sambre, prowincja Namur, Belgia) , tom 2. (red. Rougier, H. i Semal, P.) (Anthropologica et Praehistorica 124/2013, 2014).
-
Behrensmeyer, AK, Gordon, KD i Yanagi, GT Deptanie jako przyczyna uszkodzeń powierzchni kości i pseudo-nacięć. Natura 319 , 768–771 (1986).
-
Blumenschine, RJ i Selvaggio, MM Ślady uderzeniowe na powierzchniach kości jako nowa diagnostyka zachowań hominidów. Natura 333 , 763–765 (1988).
-
Pickering, TR i Egeland, CP Eksperymentalne wzorce uszkodzeń kości przez uderzenie młotkiem: implikacje dla wniosków na temat przetwarzania tusz przez ludzi. J. Archaeol. Nauka. 33 , 459–469 (2006).
-
Binford, LR Bones: starożytni ludzie i współczesne mity (Academic Press, 1981).
-
Binford, LR Nunamiut Etnoarcheologia (Academic Press, 1978).
-
Fernández-Jalvo, Y. i Andrews, P. Kiedy ludzie żują kości. J. Hum. Ewolucja 60 , 117–123 (2011).
-
Bello, SM, Parfitt, SA, De Groote, I. i Kennaway, G. Badanie eksperymentalnych uszkodzeń młotka poroża: badanie pilotażowe z wykorzystaniem obrazowania o wysokiej rozdzielczości i technik analitycznych. J. Archaeol. Nauka. 40 , 4528–4537 (2013).
-
Longin, R. Extraction du Collagène des Os Fossiles pour leur Dataation par la Méthode du Carbone 14 . Te doktoraty (Université de Lyon, 1970).
-
Mook, WG i Streurman, HJ Fizyczne i chemiczne aspekty datowania radiowęglowego. Publikacje PACT 8 , 31–55 (1983).
-
Bocherens, H. i in. Paleobiologiczne implikacje sygnatur izotopowych (13C, 15N) kopalnego kolagenu ssaków w jaskini Scladina (Sclayn, Belgia). Czwartorzędowa rez. 48 , 370–380 (1997).
-
DeNiro, MJ Konserwacja pośmiertna i zmiana stosunków izotopów kolagenu w kościach in vivo w odniesieniu do rekonstrukcji paleodietniczej. Natura 317 , 806–809 (1985).
-
Ambrose, SH Przygotowanie i charakterystyka kolagenu kostnego i zębowego do analizy izotopowej. J. Archaeol. Nauka. 17 , 431–451 (1990).
-
Rodière, E., Bocherens, H., Angibault, JM i Mariotti, A. Izotopowe właściwości azotu u saren ( Capreolus capreolus L.): implikacje dla rekonstrukcji paleośrodowiska. CR Acad. Nauka. IIa 323 , 179–185 (1996).
-
van der Plicht, J., Wijma, S., Aerts, AT, Pertuisot, MH & Meijer, HAJ Raport o stanie: Obiekt AMS w Groningen. Nukl. Instrument. Met. B 172 , 58–65 (2000).
-
Dabney, J. i in. Kompletna sekwencja genomu mitochondrialnego niedźwiedzia jaskiniowego ze środkowego plejstocenu zrekonstruowana z ultrakrótkich fragmentów DNA. Proc. Natl. Acad. Nauka. USA 110 , 15758–15763 (2013).
-
Przygotowanie biblioteki sekwencjonowania Meyer, M. i Kircher, M. Illumina do wysoce multipleksowego wychwytywania i sekwencjonowania obiektów docelowych. Harb zimnej wiosny. Protokół. 6 , pdb.prot5448 (2010).
-
Kircher, M., Sawyer, S. i Meyer, M. Podwójne indeksowanie eliminuje niedokładności w sekwencjonowaniu multipleksowym na platformie Illumina. Kwasy nukleinowe Res. 40 , e3 (2012).
-
Maricic, T., Whitten, M. i Pääbo, S. Multipleksowane przechwytywanie sekwencji DNA genomów mitochondrialnych przy użyciu produktów PCR. PLoS ONE 5 , e14004 (2010).
-
Kircher, M. W starożytnym DNA. Metody i protokoły. (red. Shapiro, B. i Hofreiter, M.) 197–228 (Humana Press, 2012).
-
Li, H. i in. Dopasowanie sekwencji/format mapy i narzędzia SAMtools. Bioinformatyka 25 , 2078–2079 (2009).
-
Li, H. i Durbin, R. Szybkie i dokładne wyrównanie krótkiego odczytu za pomocą transformacji Burrowsa-Wheelera. Bioinformatyka 25 , 1754–1760 (2009).
-
Peltzer, A. i in. EAGER: skuteczna rekonstrukcja starożytnego genomu. Biol genomu. 17 , 60 (2016).
-
Briggs, AW i in. Wzorce uszkodzeń w sekwencjach genomowego DNA neandertalczyka. Proc. Natl. Acad. Nauka. USA 104 , 14616–14621 (2007).
Podziękowanie
Dziękujemy kuratorom, którzy udostępnili nam zbiory będące pod ich opieką: A. Folie i I. Jadin (RBINS), N. Cauwe i L. Paquay (RMAH), J.-L. Schütz ( Muzeum Wielkiego Curtiusa ), B. Clarys (CHAPG) i C. Jungels ( Préhistosite de Ramioul ). Dziękujemy A. i G. Jourdan, E. Dewamme i A. Mathysowi za zdjęcia i odlewy szczątków Goyeta, I. Jadinowi za pomoc w archiwach RBINS, E. Morinowi i trzem anonimowym recenzentom za ich przydatne uwagi i sugestie . Badania te zostały sfinansowane przez Fundację Wenner-Gren (od 7837 do HR), Wyższą Szkołę Nauk Społecznych i Behawioralnych CSUN, Program wsparcia wydziału próbnego CSUN i RBINS. TEMU. był wspierany przez stypendium badawcze Marie Curie-IEF, grupę badawczą IT834-13 (Eusko Jaurlaritza/Gobierno Vasco) oraz hiszpańskie Ministerio de Economía y Competitividad (projekty CGL2012-38434-C03-01 i CGL2015-65387-C3-2 -P -MINECO/FEDER-). JK i CP otrzymali wsparcie z grantu DFG KR 4015/1-1 i Fundacji Badenii Wirtembergii.
Deklaracje etyczne
Konkurencyjne interesy
Autorzy nie deklarują żadnych konkurencyjnych interesów finansowych.
Dodatkowa informacja
Prawa i uprawnienia
Ta praca jest objęta licencją Creative Commons Uznanie autorstwa 4.0 Międzynarodowe. Obrazy lub inne materiały stron trzecich zawarte w tym artykule są objęte licencją Creative Commons artykułu, chyba że w linii kredytowej wskazano inaczej; jeśli materiał nie jest objęty licencją Creative Commons, użytkownicy będą musieli uzyskać zgodę posiadacza licencji na reprodukcję materiału. Aby wyświetlić kopię tej licencji, odwiedź http://creativecommons.org/licenses/by/4.0/
O tym artykule

Zacytuj ten artykuł
Rougier, H., Crevecoeur, I., Beauval, C. i in. Kanibalizm neandertalski i kości neandertalczyka używane jako narzędzia w Europie Północnej. Sci Rep 6 , 29005 (2016). https://doi.org/10.1038/srep29005
Raporty naukowe tom 6 , Numer artykułu: 29005 ( 2016 )
Link do artykułu: https://www.nature.com/articles/srep29005





 / Piotr Kotlarz")
{kind=link}
{kind=link}
{kind=link}