


Streszczenie
Kiła jest chorobą przenoszoną drogą płciową lub pionowo (z matki na płód), wywołaną zakażeniem Treponema pallidum subspecie pallidum (TPA). W ostatnich latach wzrosła częstość występowania kiły, mimo że bakteria ta jest obowiązkowym patogenem człowieka, droga zakażenia jest dobrze znana, a chorobę można skutecznie leczyć penicyliną. Jako środek uzupełniający kampanie zapobiegawcze i wczesne leczenie zakażonych osób, opracowanie szczepionki przeciw kiły może mieć kluczowe znaczenie w kontrolowaniu rozprzestrzeniania się i/lub nasilenia choroby, szczególnie w krajach, w których skuteczność wyżej wymienionych środków jest ograniczona. W ubiegłym stuleciu w badaniach przedklinicznych przetestowano kilka prototypów szczepionek, głównie na królikach. Chociaż żadna z nich nie zapewniała ochrony przed infekcją, niektóre prototypy zapobiegały rozprzestrzenianiu się bakterii do narządów dystalnych, osłabiały rozwój zmian chorobowych i przyspieszały ich gojenie. Pomimo tych obiecujących wyników, nadal istnieją pewne kontrowersje dotyczące identyfikacji kandydatów na szczepionki i charakterystyki odpowiedzi immunologicznej chroniącej przed kiłą. W tym przeglądzie opisujemy, co wiadomo na temat odpowiedzi immunologicznej TPA i głównych mechanizmów wykorzystywanych przez ten patogen w celu jej uniknięcia. Ponadto podkreślamy znaczenie integracji tej wiedzy w połączeniu z charakterystyką białek błony zewnętrznej (OMP) w celu przyspieszenia opracowania szczepionki przeciw kile, która może chronić przed zakażeniem TPA.
1. Wstęp
Kiła jest infekcją przenoszoną drogą płciową wywoływaną przez krętek Treponema pallidum subspecie pallidum (TPA). Choć pochodzenie epidemii kiły nie jest jasne, pierwsze przypadki tej choroby w Europie odnotowano pod koniec XV wieku ( 1 , 2 ). W 1905 roku Schaudinn i Hoffmann odkryli bakterie wywołujące tę chorobę weneryczną, a w 1943 roku pierwsze przypadki kiły udało się skutecznie wyleczyć penicyliną ( 3 ). Ponad pół wieku później penicylina pozostaje jedną z najskuteczniejszych metod leczenia ( 4 ).
Początkowo sądzono, że kiłę i trzy nieweneryczne krętkowate choroby (yaws, bejel i pinta) wywołuje ten sam czynnik, mimo że ich objawy kliniczne są różne. Ponieważ patogenne krętki są bardzo podobne morfologicznie i antygenowo (> 95% homologii DNA), konieczne było sekwencjonowanie genomu w celu zidentyfikowania odrębnych podgatunków Treponema pallidum powodujących krętki nieweneryczne: Treponema pallidum endemicum (czynnik wywołujący bejel) i Treponema pallidum pertenue (czynnik sprawczy) frambezja). Pinta jest wywoływana przez inny gatunek bakterii krętka, a mianowicie Treponema carateum ( 4 , 5 ). Wszystkie z nich są bezwzględnymi patogenami człowieka, charakteryzującymi się inwazyjnością i unikaniem odporności ( 6 – 9 ).
Zakażenie kiłą składa się z trzech głównych etapów ( 5 ). Kiła pierwotna rozpoczyna się zwykle około 2-3 tygodni po kontakcie z patogenem i charakteryzuje się obecnością owrzodzonej zmiany zwanej wrzodem. Zazwyczaj zmiana ta pojawia się w okolicy narządów płciowych lub innych częściach ciała związanych z kontaktem seksualnym, zwykle towarzyszy jej regionalna limfadenopatia ( 5 , 10 ). W przypadku braku leczenia zmiana pierwotna ustępuje samoistnie w ciągu 3-6 tygodni. Kiła wtórna rozwija się w wyniku rozprzestrzeniania się bakterii ( 10 ). Objawy kliniczne obejmują złe samopoczucie, ból głowy, gorączkę, rozsiane powiększenie węzłów chłonnych i wysypki plamisto-grudkowe z dyskretnymi lub rozległymi zmianami na całym ciele ( 5 , 10 ). Jeśli nie są leczone, zmiany wtórne mogą ustąpić nawet po kilku miesiącach. Choroba wchodzi następnie w fazę utajoną, nie wykazując objawów klinicznych ( 5 , 6 , 10 ). Nie jest jasne, w jaki sposób TPA ustala latencję i które tkanki lub narządy pełnią rolę rezerwuarów ( 6 ). Nawrót zakażenia z tych zbiorników może wystąpić u 25% nieleczonych pacjentów w ciągu dwóch lat po ustąpieniu kiły wtórnej i objawiać się nowymi, wtórnymi objawami klinicznymi ( 6 , 10 ). Po latach, a nawet dziesięcioleciach u 15–40% nieleczonych i utajonych zakażonych osób rozwinie się kiła trzeciorzędowa. Ten etap obejmuje poważne schorzenia sercowo-naczyniowe, neurologiczne, kostne i trzewne, które mogą ostatecznie doprowadzić do śmierci zakażonych osób ( 5 , 10 ). Co ciekawe, podawanie penicyliny zapobiega rozwojowi kiły do jej drugiego i trzeciego stopnia oraz leczy infekcję. Jednakże leczeni pacjenci pozostają podatni na ponowną infekcję, ponieważ poprzednia infekcja nie wywołała ochronnej odpowiedzi immunologicznej ( 11 – 13 ).
Według danych WHO w 2016 r. na kiłę chorowało 19,9 mln osób, a rocznie odnotowywano 6,3 mln nowych przypadków ( 14 ). Częstość występowania kiły wrodzonej w 2016 r. wynosiła 0,69%, przy wskaźniku 473 przypadków na 100 000 żywych urodzeń ( 15 ). Kiłę uważa się za drugą po malarii przyczynę martwych urodzeń ( 16 ). Wysoką częstość występowania kiły obserwuje się w krajach o niskich dochodach, a w ciągu ostatniej dekady liczba ta rośnie w krajach o wysokich średnich dochodach ( 5 , 14 , 17 , 18 ). W szczególności wzrost ten zaobserwowano u mężczyzn uprawiających seks z mężczyznami (MSM) z wieloma partnerami seksualnymi, a także w sieciach seksualnych osób heteroseksualnych. W tym przypadku kiła wiąże się również z większym ryzykiem zakażenia wirusem HIV ( 5 , 17 – 19 ).
Co ważne, chociaż kiłę można łatwo zdiagnozować i leczyć niedrogim antybiotykiem i jak dotąd nie zidentyfikowano żadnego rezerwuaru zwierzęcego, kiła w dalszym ciągu stanowi poważny problem zdrowotny na świecie. Dlatego pilnie potrzebne jest opracowanie szczepionki przeciw kiły, która uzupełniałaby środki kontroli i zapobiegania chorobie. W tym przeglądzie skupiamy się na: opracowaniu szczepionki przeciwko kile, zastosowaniu białek błony zewnętrznej (OMP) jako domniemanych immunogenów, odpowiedziach immunologicznych anty-TPA, a także mechanizmach unikania TPA, aby dostarczyć informacji na temat potencjalnej selekcji antygenów w przyszłych strategiach szczepionek i immunizacji .
2 Treponema pallidum subsp. blady
TPA to krętek płaskofalowy o średnicy 5-15 µm i średnicy 0,2 µm ( 20 ), należący do rodziny Spirochaetaceae, szczególnie do rodzaju Treponema. Treponemy są zwykle klasyfikowane jako bakterie Gram-ujemne ze względu na ich podwójną strukturę błonową ( 21 ). Jednak skład ich zewnętrznej błony (OM) jest zauważalnie inny ( 22 , 23 ). TPA OM charakteryzuje się niedoborem białek eksponowanych na powierzchni ( 24–26 ), obecnością fosfatydylocholiny, fosfatydyloglicerolu, fosfatydyloseryny oraz brakiem kardiolipin ( 23 ) i lipopolisacharydu (LPS) ( 27 , 28 ) . Co ciekawe, kardiolipiny są obecne w błonie cytoplazmatycznej ( 23 ) i stanowią główny antygen lipidowy, na który ukierunkowane są przeciwciała anty-TPA zakażonych osób ( 23 ). Jednakże inne składniki OM (np. glikolipidy) nie wykazują reaktywności immunologicznej ( 23 , 29 ). Skład lipidów i niska zawartość białka OM są kluczowymi cechami TPA i mogą przyczyniać się do słabej immunogenności tego patogenu ( 22 – 24 , 26 ).
TPA jest aktywnie ruchliwy, chociaż jego układ ruchowy różni się od innych bakterii wiciowych ( 30 ). Włókna osiowe (znane jako endoflagella) znajdują się w przestrzeni peryplazmatycznej i rozciągają się od biegunów komórek na całej długości ciała komórki ( 31 ). Włókna te składają się z trzech białek rdzeniowych (FlaB1, FlaB2 i FlaB3) oraz białka zewnętrznego, które otaczają rdzeń włókna (FlaA) ( 28 , 31 ). Kiedy endoflagella obraca się w jednym kierunku, ciało komórki porusza się w przeciwnym. To skręcenie powoduje ruch podobny do korkociągu, który wraz z działaniem metaloproteazy i białek adhezyny umożliwia krętkom przenikanie przez tkanki i rozprzestrzenianie się po organizmie ( 32 , 33 ).
Do niedawna TPA nie hodowano in vitro ( 34 ), a namnażanie bakterii u królików było jedyną strategią pozwalającą na uzyskanie wystarczającej liczby żywych i zakaźnych organizmów do badań eksperymentalnych ( 35 ). Brak cyklu kwasów trikarboksylowych i wymagania mikroaerofilne sprawiają, że bakteria ta jest całkowicie zależna od komórek gospodarza w zakresie pozyskiwania puryn, pirymidyn i większości aminokwasów ( 6 , 8 , 28 ). Dlatego TPA nie może przetrwać poza żywicielem i traci zdolność zakaźną w ciągu kilku godzin ( 8 , 36 ).
Oprócz ludzi króliki są jednymi z niewielu ssaków podatnych na infekcję TPA i stały się zwierzęcym modelem referencyjnym do badań ochrony immunologicznej przed kiłą. Po zakażeniu u królików rozwijają się objawy kliniczne pierwotne i wtórne oraz reakcja humoralna podobna do tej obserwowanej u ludzi ( 37 – 39 ). Śródskórna inokulacja TPA powoduje zmiany skórne przypominające ludzkie wrzody, a bakterie mogą rozprzestrzenić się do narządów dystalnych, głównie wtórnych narządów limfatycznych (tj. śledziony) ( 32 , 40 ). Jednakże inwazja na ośrodkowy układ nerwowy nie jest często obserwowana i zależy to od użytego szczepu TPA ( 41 ). Oprócz królików istnieją również inne gatunki podatne na TPA (tj. naczelne inne niż ludzie, chomiki, świnki morskie i myszy). Chociaż objawy kliniczne podobne do ludzkich występują jedynie u naczelnych innych niż ludzie i królikach ( 40 , 42 ), myszy i inne gryzonie mogą być przydatne do badania rozprzestrzeniania się bakterii ( 43 ).
Genom TPA jest kolistym chromosomem o około 1138 kilopar zasad, który zawiera 1041 przewidywanych ORF ( 28 ). Jej genom jest mały w porównaniu z innymi bakteriami chorobotwórczymi ( 28 ). Sekwencja genomu potwierdza, że TPA nie może syntetyzować de novo kofaktorów enzymów, kwasów tłuszczowych, enzymów szlaku trikarboksylowego ani nukleotydów, mimo że zawiera 57 ORF kodujących białka transportowe i jest w stanie wykorzystywać szlak glikolityczny ( 28 ). TPA brakuje genów kodujących dysmutazę ponadtlenkową, katalazę lub peroksydazę, co mogłoby wyjaśniać jego podatność na tlen. Dodatkowo zidentyfikowano także kilka genów zaangażowanych w syntezę białek ruchliwych i lipoprotein ( 4 , 8 , 28 ). Porównanie genomów patogennych podgatunków Treponema pallidum wykazało, że są one podobne pod względem wielkości i struktury, a różnią się sekwencją genomu o mniej niż 0,2–0,4% ( 44 , 45 ).
3 Przebieg zakażenia i odpowiedź immunologiczna
Kiła jest zazwyczaj przenoszona poprzez kontakt seksualny lub z matki na dziecko. Krętki uzyskują dostęp do żywiciela poprzez mikrootarcia naskórka lub bezpośrednio penetrujące błony śluzowe ( 46 , 47 ). Gdy TPA dostanie się do organizmu, przylega do komórek nabłonkowych i macierzy zewnątrzkomórkowej i lokalnie namnaża się z szacunkową szybkością raz na 30–33 godziny ( 48 , 49 ). Badania wiązania in vitro wykazały, że laminina i fibronektyna należą do cząsteczek kotwicowych biorących udział w tych interakcjach ( 50 – 53 ). Ponieważ TPA nie zawiera toksyn cytotoksycznych ani innych czynników zjadliwości ( 28 ), zniszczenie tkanki i zmiany wrzodowe są prawdopodobnie spowodowane reakcją zapalną w miejscu wniknięcia ( 47 ). Początkowa odpowiedź immunologiczna usuwa bakterie miejscowo, a zmiany w stadium pierwotnym ustępują samoistnie w ciągu 3–6 tygodni. W międzyczasie krętki rozprzestrzeniają się po całym organizmie, a infekcja szybko staje się ogólnoustrojowa, wywołując kiłę wtórną. Następnie krętki penetrują głębsze tkanki i indukują ekspresję sygnałów zapalnych, promując migrację komórek odpornościowych do zakażonych tkanek ( 47 ). Badania in vitro wykazały, że TPA może indukować ekspresję cząsteczek adhezyjnych ICAM-1, VCAM-1 i E-selektyny w komórkach śródbłonka ( 54 – 56 ). Ponadto aktywność metaloproteazy odgrywa główną rolę w penetracji i rozprzestrzenianiu się TPA przez macierz pozakomórkową i połączenia międzykomórkowe ( 57 , 58 ). Na ostatnim etapie infekcja i związana z nią odpowiedź immunologiczna uszkadzają różne narządy (np. serce i mózg), powodując wyniszczające problemy zdrowotne ( 47 ).
Zakażenie TPA wyzwala złożoną odpowiedź immunologiczną, która nie kontroluje rozprzestrzeniania się bakterii i postępu choroby. W kile pierwotnej leukocyty wielojądrzaste (PMN) są pierwszymi komórkami odpornościowymi, które naciekają miejsce zakażenia ( 59 ) ( ryc. 1A ). Komórki te mogą przyczyniać się do początkowej kontroli infekcji poprzez wydzielanie peptydów przeciwdrobnoustrojowych ( 60 ) i usuwanie krętków na drodze fagocytozy ( 59 , 61 ). Jednak początkowa kontrola infekcji jest ograniczona, ponieważ praktycznie we wszystkich przypadkach dochodzi do rozprzestrzeniania się bakterii. Oprócz PMN, komórki dendrytyczne (DC) ( Figura 1A ) mogą również fagocytować całe fragmenty lub fragmenty pochodzące od bakterii, stymulując w ten sposób ich dojrzewanie i zdolność do prezentacji antygenu. Zatem DC zwiększają wytwarzanie cytokin prozapalnych, takich jak IL-12, IL-6 i IL-1B, i regulują w górę ekspresję CD54, CD83, CD80, CD86 i HLA-DR (62 , 63 ) . Dojrzałe DC migrują do węzłów chłonnych, gdzie prezentują antygeny krętkowe limfocytom T, indukując odpowiedź limfocytów T specyficzną dla antygenu. Te limfocyty T można wykryć we wtórnych narządach limfatycznych zakażonych zwierząt trzy dni po zakażeniu i stopniowo gromadzą się w miejscu zakażenia ( 64 – 66 ), co zbiega się z maksymalnym obciążeniem TPA ( 67 , 68 ). Króliki zakażone TPA wykazywały szybki rozrost stref limfocytów T we wtórnych narządach limfatycznych (węzłach chłonnych i śledzionie) ( 38 , 66 ). Chociaż w nacieku limfocytów T dominują limfocyty Th1 CD4 + ( 38 , 61 , 69 ) ( Figura 1A ), cytotoksyczne limfocyty T (CD8 + ) można znaleźć także w zmianach pierwotnych, gdzie ich rola pozostaje niejasna. Komórki T CD8 + mogą być wymagane do eliminacji rezerwuaru TPA wewnątrz komórek niefagocytarnych we wczesnych zmianach kiłowych ( 69 – 71 ). W tym sensie perforynę i granzym wykrywa się we wczesnych zmianach chorobowych, co sugeruje aktywność cytolityczną ( 72 ). Co więcej, komórki T CD8 + , jak również komórki NK, mogą przyczyniać się do wydzielania interferonu-γ (IFN-γ) ( 9 , 73 ). Cytokiny Th1 mogą promować migrację i aktywację makrofagów ( Figura 1A ), których liczba gwałtownie wzrasta do 10 dnia po zakażeniu w miejscu wejścia ( 68 ). Aktywowane makrofagi fagocytują i niszczą krętki ( 64 , 74) do czasu całkowitego usunięcia ich z miejsca zakażenia. Co ciekawe, proces ten może zostać wzmocniony przez przeciwciała opsonizujące TPA i IFN-γ, szczególnie poprzez interakcję z receptorem Fc-Fcγ ( 73 – 77 ). Jednakże inne, mniej wydajne mechanizmy, takie jak fagocytoza nieopsoniczna i aktywna bezpośrednia inwazja, mogą również uczestniczyć w wzajemnym oddziaływaniu TPA-makrofagi ( 78 ). W odpowiedzi na zakażenie TPA makrofagi polaryzują się w kierunku fenotypu M1 i wydzielają cytokiny prozapalne (tj. IL-1β, TNF-α, IL-12 i IL-15) (77, 79), które mogą ostatecznie sprzyjać martwicy i tworzeniu się wrzodów typowych dla pierwotne zmiany wrzodowe.
Rysunek 1
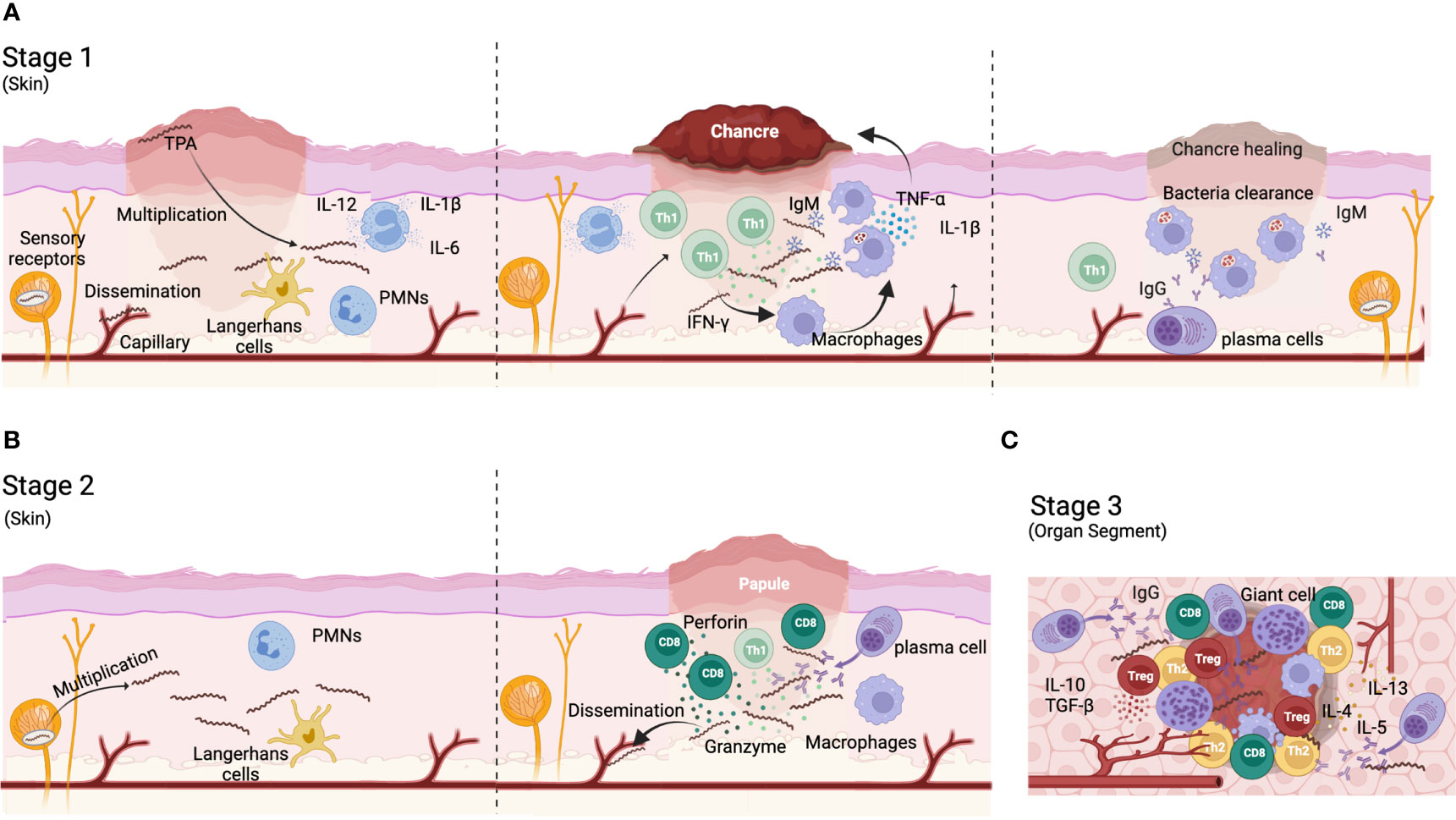
Jeśli chodzi o odpowiedź humoralną, przeciwciała anty-TPA można wykryć już 6 dni po zakażeniu ( 80 ). Jednakże kinetyka odpowiedzi przeciwciał różni się w zależności od docelowego białka i szczepu TPA. Na przykład, poziom przeciwciał skierowanych przeciwko podrodzinie I i II Tpr wzrasta pomiędzy 10. a 45. dniem po zakażeniu ( 81 ). Podczas pierwotnej infekcji w odpowiedzi humoralnej anty-TPA dominuje IgM, podczas gdy poziom IgG stopniowo wzrasta ( 80 , 82 ) ( Figura 1A ). Przeciwciała biorą udział nie tylko w procesach opsonizacji, ale mogą również blokować rozprzestrzenianie się bakterii i powstawanie zmian skórnych. Co więcej, układ dopełniacza jest również zaangażowany w ograniczanie aktywności krętków, szczególnie współpracując z przeciwciałami ( 83 – 85 ). TPA gromadzi kwas sialowy na swojej powierzchni, czyniąc go odpornym na lizę dopełniacza drogą alternatywną ( 86 – 89 ). Zatem klasyczny szlak aktywacji dopełniacza jest kluczem do usuwania bakterii. Dane te sugerują, że odpowiedź immunologiczna generowana przeciwko TPA podczas pierwotnej infekcji jest odpowiedzią immunologiczną związaną z nadwrażliwością typu opóźnionego (DTH), w której główną rolę odgrywają uczulone limfocyty T. W rzeczywistości postawiono hipotezę, że rokowanie w przypadku kiły zależy od równowagi pomiędzy DTH a reakcjami humoralnymi. W związku z tym zaproponowano, że do usunięcia infekcji może być wymagana silna odpowiedź immunologiczna na DTH, podczas gdy opóźnienie może wynikać z pośrednich odpowiedzi DTH. Choroba trzeciorzędowa będzie związana ze słabym DTH i silną odpowiedzią humoralną ( 68 , 90 ). Mimo to przeciwciała mogą mieć kluczowe znaczenie w zapobieganiu infekcji ( 68 , 91 ). W związku z tym u królików otrzymujących surowicę odpornościową wykazano opóźnienie w pojawianiu się zmian chorobowych, które były również mniej poważne ( 92 ). Co więcej, pacjenci, którzy byli wcześniej leczeni na kiłę, pozostają podatni na ponowne zakażenie, ponieważ nie rozwija się u nich skuteczna odpowiedź humoralna anty-TPA [ 68 ].
Zarówno odpowiedź komórkowa, jak i humoralna utrzymują się przez miesiące po ustąpieniu pierwotnych zmian kiłowych ( 39 , 93 , 94 ), a ich nasilenie koreluje z utrzymywaniem się TPA jako zakażenia utajonego ( 95 – 98 ). Od tego stanu może nastąpić nawrót aktywnej infekcji, prowadzący do kiły wtórnej i trzeciorzędowej. Kiła wtórna objawia się zapaleniem skóry, które różni się wyglądem histologicznym od kiły pierwotnej. Histologia kiły wtórnej jest zmienna, ale ogólnie rzecz biorąc, w zmianach wtórnych często występują limfocyty, makrofagi i komórki plazmatyczne, podczas gdy nacieki wielojądrzaste i eozynofilowe występują w mniejszym odsetku (99–101) (ryc . 1B ) . Co więcej, limfocyty T CD8 + są licznie reprezentowane ( Figura 1B ) ( 99 ). W przeciwieństwie do kiły pierwotnej, w przypadku kiły wtórnej w humoralnej odpowiedzi immunologicznej anty-TPA dominują przeciwciała IgG ( ryc. 1B ) ( 82 , 102 , 103 ). Co więcej, podczas gdy IgG1 jest główną podklasą IgG w kile pierwotnej, zarówno podklasy IgG1, jak i IgG3 można w równym stopniu znaleźć w kile wtórnej ( 104 ).
Wreszcie kiła trzeciorzędowa charakteryzuje się uszkodzeniem wielu narządów i rozwojem dziąseł w różnych tkankach (takich jak wątroba i skóra). Gumy to struktury przypominające ziarniniaki, charakteryzujące się nekrotycznym jądrem otoczonym makrofagami, gigantycznymi komórkami wielojądrowymi, limfocytami i komórkami plazmatycznymi ( 68 ) ( Ryc. 1C ). Kiła układu nerwowego charakteryzuje się wzrostem liczby limfocytów T CD8 + we krwi ( 105 ). Ponadto w trakcie ewolucji choroby obserwuje się zmianę wytwarzania cytokin Th1 na Th2. Podczas gdy cytokiny Th1 są wykrywane głównie podczas kiły pierwotnej, stężenie cytokin Th2 wzrasta w późnych stanach choroby ( 106 ) ( Ryc. 1C ). Koreluje to ze zmniejszeniem liczby komórek w rozproszonej korze i rozrostem pęcherzykowym w węzłach chłonnych z akumulacją komórek plazmatycznych w obszarach międzypęcherzykowych podczas kiły trzeciorzędowej ( 68 ). Te zmiany w odpowiedzi immunologicznej podczas kiły są ściśle powiązane z postępem choroby.
4 Badania nad immunizacją i strategia szczepień
Pomimo kampanii na rzecz zdrowia publicznego i dostępności skutecznego leczenia, w ciągu ostatniej dekady częstość występowania kiły na całym świecie wzrosła ( 17 ), co sugeruje potrzebę wprowadzenia dodatkowych środków w celu kontroli przenoszenia infekcji. Cennym narzędziem może być opracowanie szczepionki na kiłę. Jednak po kilkudziesięciu latach badań skuteczna szczepionka na kiłę pozostaje nieuchwytna. Przetestowano różne strategie, w tym inaktywowane bakterie i rekombinowane białka podjednostkowe, chociaż z ograniczonym sukcesem. Co ciekawe, Miller i wsp. wykazali ochronę królików przed zakażeniem TPA po 60 immunizacjach bakteriami napromieniowanymi promieniami γ przez 37 tygodni ( 107 ). Chociaż ta strategia eksperymentalna jest daleka od zastosowania u ludzi, służy jako dowód na to, że szczepionka na kiłę jest wykonalna. Ponadto badania prowokacyjne u ludzi przeprowadzone w latach 50. wykazały, że pacjenci zakażeni w sposób utajony byli odporni na ponowne zakażenie heterologicznym szczepem TPA ( 108 ). W związku z tym Marra i wsp. podali, że wcześniejsze zakażenie kiłą może złagodzić objawy późniejszego zakażenia TPA ( 109 ). Zatem u osób, które przeżyły trzy lub więcej epizodów kiły, ryzyko rozwoju utajonej kiły wczesnej było większe po kolejnych infekcjach. Badania te wskazują, że wytworzenie ochronnej odporności przeciwko TPA zajmie dużo czasu, co dodatkowo podkreśla niską immunogenność tego patogenu.
Do 2018 r. jedną z głównych przeszkód w opracowywaniu szczepionki przeciwko kiły był brak możliwości hodowli bakterii in vitro ( 34 ). To ograniczenie techniczne przesunęło punkt ciężkości większości badań w stronę rekombinowanych OMP ( 110 ). Chociaż żadne z tych białek nie zapewniało całkowitej ochrony przed prowokacją TPA in vivo , zaobserwowano obiecujące wyniki; mianowicie wywołanie silnej odpowiedzi humoralnej, zmniejszenie zmian wrzodziejących, szybsze gojenie się zmian chorobowych lub zahamowanie rozprzestrzeniania się bakterii do narządów dalszych. Badania te dostarczyły znaczącej wiedzy na temat OMP, schematów szczepień i potencjalnych celów szczepionek. Postęp narzędzi bioinformatycznych umożliwił identyfikację dodatkowych przypuszczalnych OMP, a także przewidywanie modeli struktury i epitopów komórek B ( 111 , 112 ). Zastosowanie nowo opracowanych podejść (np. inżynierii genetycznej) i udoskonalonych narzędzi bioinformatycznych umożliwia dalsze zgłębianie wiedzy na temat OMP, co przyniesie korzyści przyszłym badaniom nad szczepionkami ( 113 , 114 ).
5 Białka błony zewnętrznej
Jedną z głównych cech odróżniających TPA od innych bakterii jest niedostatek OMP eksponowanych na powierzchni ( 24 – 26 ). Obliczono, że gęstość OMP w TPA jest w przybliżeniu 100-krotnie mniejsza niż w E. coli ( 22 ). Identyfikacja i charakterystyka OMP TPA była wyzwaniem ze względu na niemożność manipulacji genetycznej i hodowli TPA in vitro (do niedawna) ( 34 ) oraz kruchość jego OM, co wywołało silne kontrowersje w niektórych badaniach. Wewnętrzne właściwości OM (tj. brak LPS i mała gęstość białek) sprawiają, że można go łatwo uszkodzić w wyniku typowych manipulacji eksperymentalnych (np. wirowania, ponownego zawieszania lub stosowania niejonowego detergentu w niskim stężeniu) ( 22 , 26 ) . Zakłócenie OM prowadzi do potencjalnej ekspozycji lipoprotein, które normalnie są obecne w przestrzeni peryplazmatycznej i błonie cytoplazmatycznej, i może błędnie wskazać białka jako OMP. Ze względu na te ograniczenia techniczne identyfikacja OMP często opierała się na przewidywaniu ich sekwencji i struktury lub na porównaniu z funkcjonalnymi ortologami innych gatunków ( 111 ). Zatem OMP można podzielić na trzy główne grupy według ich domniemanych funkcji:
5.1 Funkcja transportowa
Ponieważ import składników odżywczych i wydalanie substancji toksycznych mają kluczowe znaczenie dla nadzoru nad bakteriami, w funkcję transportu zaangażowanych jest kilka OMP TPA.
5.1.1 Rodzina białek Tpr
Rodzina białek powtórzeń Treponema pallidum (Tpr) obejmuje 12 członków, którzy są podzieleni na trzy podrodziny w zależności od składu aminokwasów: podrodzina I (TprC, D, F, I), podrodzina II (TprE, G, J) i podrodzina III (TprA, B, H, K, L). Wśród nich najszerzej badane są podrodzina I i TprK (podrodzina III). Jeśli chodzi o lokalizację białka Tpr, analiza sekwencji przewidywała, że TprB, TprC, TprD, TprE, TprG, TprH, TprI, TprJ, TprK i TprL mogą być zlokalizowane w OM (22 ) . Na poparcie tej przewidywania zidentyfikowano przypuszczalny rozszczepialny peptyd sygnałowy u większości członków nadrodziny Tprs, w tym białek TprC, D, F, I, E, G, J, A, B i TprK (111 , 115 ) .
Rodzina Tpr jest spokrewniona z głównym białkiem osłonki zewnętrznej (MOSP) Treponema denticola ( 115 ), białkiem eksponowanym na powierzchni, posiadającym funkcję adhezyjną i porinową ( 116 – 118 ). Większość Tpr obejmuje trzy główne domeny: 1) domenę N-końcową związaną z domeną N-końcową Treponema denticola MOSP, 2) region centralny i 3) domenę C-końcową związaną z domeną C-końcową Treponema dentiola MOSP, któremu brakuje białek TprF i TprA. Ostatnio przewidywano obecność krótkich cech rozpoznawania molekularnego (MoRF) w większości Tpr. Motywy te mogą oddziaływać ze specyficznymi białkami i po związaniu przechodzić od zaburzenia do porządku ( 111 ). Analiza sekwencji wykazała, że domeny N- i C-końcowe białek Tpr z podrodziny I i II są stosunkowo konserwatywne, podczas gdy domeny centralne mają zmienną długość i sekwencję ( 6 ). Domeny końcowe przedstawiają regiony amfipatyczne i flankujące regiony hydrofilowe domen centralnych. Według Hawley i wsp. domeny N- i C-końcowe ze wszystkich Tprs przyjmują strukturę β-niciową, podczas gdy regiony centralne mogą być głównie α-helisami ( 111 ).
Analiza strukturalna podrodziny I wykazała, że białka te tworzą trimeryczny kanał β-beczułki. Jednakże w przeciwieństwie do klasycznych porin, których cały polipeptyd tworzy beczkę β, podrodzina Tpr I posiada topologię dwudzielną. Domena C-końcowa tworzy strukturę beczki β i jest wystawiona na powierzchnię, podczas gdy domeny N-końcowa i środkowa zakotwiczają beczkę β w worku peptydoglikanowym obszaru peryplazmatycznego ( 119 , 120 ). Odpowiednio, chociaż ekspresja TprC/D w E. coli jest eksponowana na powierzchni, TprF nie posiada domeny C-końcowej i dlatego jest całkowicie periplazmatyczna ( 22 ). Immunodetekcja białek Tpr za pomocą mikroskopii elektronowej przy użyciu oczyszczonych króliczych immunoglobulin anty-Tpr I potwierdziła obecność białek Tpr w OM i przestrzeniach peryplazmatycznych, co może wynikać z opisanej wcześniej struktury dwudzielnej lub z białek rozpoznawanych przez przeciwciało anty-Tpr I z odrębna lokalizacja ( 121 ). Niedawno Hawley i wsp. potwierdzili, że topologia dwuczęściowej błony zidentyfikowana w podrodzinie I jest wspólna dla wszystkich Tpr z trzema pełnymi domenami ( 111 ). Na podstawie analizy sekwencji i struktury wykazano, że rodzina Tpr może brać udział w imporcie małych rozpuszczalnych cząsteczek. W tym sensie integracja niektórych białek Tpr w liposomach zwiększyła ich przepuszczalność ( 119 , 120 ). Dlatego zmiany w ekspresji białek i/lub zmienność ich regionu β-beczki tworzącego kanały można wykorzystać do dostosowania wymagań żywieniowych TPA do środowiska. W związku z tym ekspresja białek Tpr może różnić się pomiędzy szczepami, a także w tym samym szczepie w czasie ( 122 ), co może wyjaśniać, dlaczego odpowiedź humoralna anty-Tpr różni się między izolatami TPA ( 81 ). Obserwacja ta sugeruje istnienie mechanizmów, które mogą regulować ekspresję tych genów. W ten sposób zidentyfikowano hiperzmienny homopolimeryczny szlak guanozyny (poli-G) i motyw wiążący białko receptora cAMP (CRP) powyżej miejsca startu transkrypcji podrodziny Tpr II i III ( 123 – 125 ). W rzeczywistości Tp0262 (homolog TPA CRP) może wiązać się z tymi promotorami i regulować ekspresję powierzchniową tych białek podczas infekcji ( 123 ). W rezultacie specyficzność odpowiedzi humoralnej wywołanej wobec białek Tpr różni się w zależności od izolatu TPA ( 81 ).
Z nadrodziny Tpr, TprK jest jednym z najlepiej scharakteryzowanych białek. Wykazano wcześniej, że rekombinowany TprK jest monomeryczną poriną, metodą barwienia negatywowego pod mikroskopem elektronowym ( 126 ). Przeciwciała anty-TprK można wykryć już w trakcie infekcji [ 81 ]. Jednakże TprK wykazuje zmienność sekwencji, która przyczynia się do ucieczki odporności. Zatem zidentyfikowano kilka alleli wśród szczepów TPA ( 127 , 128 ). Co więcej, białko to wykazuje siedem domen zmiennych, których sekwencja zmienia się w wyniku rekombinacji niewzajemnej, głównie z regionami położonymi poniżej genu TprD ( 127 , 128 ), a mechanizm ten nasila się pod presją immunologiczną ( 129 , 130 ). Istnienie tego mechanizmu może wyjaśniać, dlaczego zmienność TprK była większa w izolatach TPA od pacjentów chorych na kiłę wtórną niż u pacjentów z kiłą pierwotną ( 128 ). Jednakże zmienność genetyczna TprK może również wystąpić przy braku presji immunologicznej ( 131 ). Co ciekawe, donoszono, że odpowiedź humoralna in vivo jest ukierunkowana na regiony zmienne, podczas gdy odpowiedź komórkowa rozpoznaje konserwatywne epitopy TprK ( 132 , 133 ). Co więcej, zwierzęta prowokowane przy użyciu TPA z TprK homologicznym do tego stosowanego w schemacie immunizacji wykazują lepszą ochronę niż zwierzęta prowokowane szczepem wyrażającym heterologiczny TprK ( 132 ). Mimo to istnieją kontrowersje dotyczące roli przeciwciał anty-TprK, skuteczności ochronnej TprK jako immunogenu i jego lokalizacji. Centurion-Lara i współpracownicy opisali, że TprK jest zlokalizowany na OM, a przeciwciała specyficzne dla TprK mają zdolność opsonizacji ( 115 ). Co ciekawe, u królików immunizowanych tym białkiem nie rozwinęły się zmiany wrzodziejące. Zmiany te goiły się szybciej niż te obserwowane u nieszczepionych zwierząt kontrolnych i zawierały mniej krętków, gdy analizowano je pod mikroskopem w ciemnym polu ( 115 ). Dodatkowo Morgan i in. zidentyfikowali region N-końcowy TprK (37-273 aminokwasów) i, w mniejszym stopniu, część C-końcową (349-478 aminokwasów) jako części białka, które indukowały opisaną wcześniej ochronną odpowiedź immunologiczną ( 134 ). Z drugiej strony Hazlett i wsp. opisali, że TprK był zlokalizowany w przestrzeni peryplazmatycznej, przeciwciała anty-TprK nie wykazywały aktywności opsonizacyjnej, a immunizacja rekombinowanym TprK nie indukowała ochronnej odpowiedzi immunologicznej ani zmienności sekwencji TprK (135 ) . Aby wyjaśnić te rozbieżności, konieczne byłoby dalsze badanie.
Jeśli chodzi o odporność wywołaną przeciwko innym członkom Tprs, u 98% osób chorych na kiłę wykryto przeciwciała anty-TprI. Przeciwciała te atakowały głównie konserwatywny region N-końcowy. Brak reaktywności powiązano z wczesną infekcją ( 121 ). Co ciekawe, eksperymenty przeprowadzone na królikach wykazały, że immunizacja TprI nie chroniła królików przed infekcją ( 136 ). Jednakże zmiany skórne nie ulegały owrzodzeniu i goiły się szybciej niż u nieszczepionych zwierząt. Białko to wywoływało silną odpowiedź humoralną i komórkową, ukierunkowaną głównie na konserwatywny region N-końcowy białka podczas zakażenia TPA ( 81 , 136 ). Nic dziwnego, że immunizacja konserwatywnym regionem N-końcowym TrpF, który jest również wspólny dla wszystkich członków podrodziny I, nie chroniła królików przed zakażeniem TPA po prowokacji. Jednakże szczepienie TprF osłabiło rozwój zmian chorobowych, zapobiegło owrzodzeniom i zmniejszyło obciążenie bakteryjne w zmianach skórnych ( 136 ). W odniesieniu do TprC i TprD mapowanie epitopów wykazało, że odpowiedź humoralna wykryta w surowicy królika jest skierowana przeciwko odsłoniętym pętlom ( 112 ). Co więcej, regiony N- i C-końcowe zawierają najbardziej reaktywne epitopy. Co zaskakujące, Anand i wsp. wykazali, że surowice od zakażonych osób nie rozpoznają TprC, podczas gdy króliki zakażone TPA wytwarzają przeciwciała o działaniu opsonizacyjnym ( 119 ). Jednym z wyjaśnień może być niska ekspresja TprC i różna wielkość odpowiedzi humoralnej stwierdzana u różnych szczepów i podgatunków kiły.
5.1.2 Tp0515
Tp0515 jest strukturalnym ortologiem białka LptD, które jest częścią kompleksu wielobiałkowego zaangażowanego w transport LPS do OM u bakterii Gram-ujemnych ( 22 ). Zatem, ponieważ LptD jest osadzony w OM, wnioskuje się, że Tp0515 jest OMP. Jednakże analiza sekwencji genomu TPA nie pozwoliła zidentyfikować szlaku biosyntezy LPS ( 28 ). Warto zauważyć, że w TPA odkryto białka ortologiczne dla kilku składników szlaku transportu LPS ( 111 ), co sugeruje, że nawet bez LPS białka te mogłyby brać udział w translokacji innych produktów komórkowych do OM, takich jak glikolipidy ( 22 ). Modelowanie strukturalne Tp0515 wykazało, że białko to jest 26-niciową beczką β osadzoną w OM z odsłoniętymi pętlami zewnątrzkomórkowymi ( 111 ). Co więcej, Hawley i in. zidentyfikowali epitopy komórek B wewnątrz tych pętli zewnątrzkomórkowych za pomocą analizy bioinformatycznej ( 111 ). Jednakże immunogenność i zdolność tego białka do wywoływania ochronnej odpowiedzi immunologicznej nadal wymagają empirycznego wykazania.
5.1.3 Tp0126 i inne ortologi OmpW/AlkL
W TPA zidentyfikowano cztery białka będące ortologami OmpW (Tp0126 i Tp0733) i OprG (Tp0479 i Tp0698) ( 111 ). Zarówno białka E. coli OmpW, jak i Pseudomonas aeruginosa OprG należą do rodziny białek OmpW/AlkL, co do których sugeruje się, że biorą udział w adaptacji bakterii do stresu środowiskowego i pozyskiwaniu składników odżywczych ( 137 , 138 ). OmpW i OprG mają ośmioniciową architekturę β-beczułki i tworzą swoisty kanał hydrofobowy, który umożliwia transport cząsteczek hydrofobowych bezpośrednio do membrany z pominięciem hydrofilowej przestrzeni peryplazmatycznej ( 111 , 139 ) . Zatem Tp0126 może brać udział w transporcie kwasów tłuszczowych (nie może być syntetyzowany przez sam TPA) ( 140 ). Co ciekawe, białko to jest wysoce konserwatywne wśród szczepów TPA. Jego transkrypcja jest regulowana przez obecność homopolimerów guanidyny o różnej długości, zlokalizowanych przed promotorem, co jest zgodne ze zmiennością fazową ( 141 ).
Co ciekawe, Tp0126 może wykazywać niską immunogenność. Poziomy przeciwciał anty-Tp0126 w próbkach surowicy od pacjentów z kiłą utajoną były niższe niż te skierowane przeciwko Tp0574, wysoce wyrażanemu i immunogennemu białku TPA ( 141 ). Ponadto przeciwciała anty-Tp0126 wykryto u królików immunizowanych Tp0126 dopiero po trzeciej dawce przypominającej, co podkreśla potencjalnie niską immunogenność tego białka ( 141 ). Co ciekawe, przeciwciała te celowały w pętle zewnętrzne Tp0126 i były zdolne do opsonizacji TPA. Jednakże immunizacja Tp0126 wykazała słabą zdolność ochronną, jeśli w ogóle, u królików. W rzeczywistości izolaty krętków ze zmian chorobowych u immunizowanych i prowokowanych królików wykazywały niższy poziom transkrypcji Tp0126, co było związane z dłuższym regionem poliG (≥ 9 G), w porównaniu z tremponemami uzyskanymi z inokulum prowokacyjnego (141 ) . Wyniki te sugerują, że ekspresja Tp0126 może być zmniejszona podczas infekcji, co może wyjaśniać brak ochrony obserwowany w eksperymentach immunizacji i prowokacji.
Przewiduje się , że podobnie jak Tp0126, inni członkowie grupy ortologów TPA OmpW/AlkL (Tp0733, Tp0479 i Tp0698) ( 111 ) utworzą kanał błonowy i będą zaangażowani w transport małych cząsteczek. Hawley i in. przewidywali wiele eksponowanych na powierzchni epitopów komórek B w ich pętlach zewnątrzkomórkowych, chociaż gęstość tych epitopów różni się w zależności od białka ( 111 ). Przewiduje się, że przyszłe badania eksperymentalne z tymi OMP potwierdzą ich przewidywania in silico oraz pozwolą uzyskać informacje na temat potencjalnej immunogenności i zdolności do wywoływania ochronnych odpowiedzi immunologicznych.
5.1.4 Białka podobne do FadL
Ze względu na niską przepuszczalność OM bakterii Gram-ujemnych, bakterie te posiadają maszynerię białkową OM zaangażowaną w wychwyt długołańcuchowych kwasów tłuszczowych, taką jak FadL (białko transportujące długołańcuchowe kwasy tłuszczowe). TPA nie jest w stanie syntetyzować długołańcuchowych kwasów tłuszczowych ( 28 ). Ponieważ jednak TPA OM jest bardziej przepuszczalny dla kwasów tłuszczowych niż inne bakterie Gram-ujemne ( 142 ), dyfuzja błonowa może być jednym z mechanizmów wykorzystywanych przez ten mikroorganizm do wychwytu tych cząsteczek. Niemniej jednak w TPA zidentyfikowano pięć ortologów FadL (Tp0548, Tp0856, Tp0858, Tp0859 i Tp0865), co sugeruje, że bierna dyfuzja kwasów tłuszczowych nie jest jedynym mechanizmem (111 ) . Modelowanie strukturalne tych ortologów przewidywało, że wszystkie białka podobne do TPA FadL tworzą 14-niciową beczkę β z N-końcem w świetle beczki ( 111 ). Co więcej, wszystkie pięć białek zawiera jeden lub więcej przewidywanych epitopów dla limfocytów B ( 111 ). Co ciekawe, Delgado i in. niedawno opisali obecność przeciwciał IgG i komórek IgG + B u królików zakażonych TPA, które rozpoznały pętle zewnątrzkomórkowe 2 i 4 z Tp0856 i Tp0858 ( 143 ), wskazując, że białka te mogą być przydatne w projektowaniu szczepionek.
5.1.5 Białka podobne do TolC
TolC to białko OM z E. coli , które jest częścią kompleksu pompy wypływowej zaangażowanej w usuwanie substancji toksycznych ( 144 ). Do chwili obecnej w TPA opisano cztery ortologi TolC (Tp0966, Tp0967, Tp0968 i Tp0969) ( 145 ). Modelowanie strukturalne tych białek wykazało, że mają one topologię podobną do TolC opartą na czterech niciach β z dwiema dużymi pętlami zewnątrzkomórkowymi i sześcioma α-helisami ( 111 ). Ponadto Hawley i in. przewidzieli, że wszystkie cztery białka TPA podobne do TolC, w szczególności Tp0969, otaczają epitopy komórek B wystawione na powierzchnię ( 111 ). Zatem te przewidywane OMP mogą być celem układu odpornościowego. Jednakże konieczna jest dalsza analiza ich immunogenności in vivo w celu określenia ich potencjału w opracowywaniu szczepionek.
5.1.6 Rzadkie białka błony zewnętrznej krętka (TROMP)
Rodzina TROMP obejmuje trzy białka: TROMP-1 (31 kDa), TROMP-2 (28 kDa) i TROMP-3 (65 kDa) ( 146 ). TROMP-1 (zwany także TroA lub Tp0163) został po raz pierwszy opisany jako białko eksponowane na powierzchni o właściwościach przypominających porinę ( 147 ). Jednakże później zidentyfikowano, że jest to metaloproteina peryplazmatyczna zakotwiczona w błonie cytoplazmatycznej ( 148 – 150 ). TROMP-2 (zwany także Tp0663) zlokalizowano na OM, gdy ulegał ekspresji jako białko rekombinowane w E. coli . Jednakże izolacja TROMP-2 z E. coli OM wykazała niewielką ilość insercji poryn, co budzi wątpliwości co do jej funkcji ( 29 ). Co ciekawe, zarówno TROMP-1, jak i TROMP-2 były celem przeciwciał obecnych w surowicy zakażonych królików i ludzi ( 146 ). W rzeczywistości TROMP-2 mógłby być potencjalnym kandydatem do serodiagnostyki wszystkich stadiów kiły ( 151 ). Doświadczenia z prowokacją TPA przeprowadzone na królikach immunizowanych Tp0663 wykazały częściową ochronę przy wysokich mianach przeciwciał anty-Tp0663. Autorzy zaobserwowali osłabienie zmian chorobowych ze wzrostem nacieku komórkowego. Co ważne, u immunizowanych zwierząt nie wykryto rozprzestrzeniania się bakterii do narządów dalszych ( 152 ). Warto zauważyć, że TROMP-3 ulega ekspresji w niższym stężeniu niż pozostali dwaj członkowie tej rodziny, a jego funkcja i struktura pozostają nieznane ( 29 ).
5.2 Funkcja przyczepności
Adhezja do komórek i macierzy zewnątrzkomórkowej ma kluczowe znaczenie dla wywołania zakażenia i rozprzestrzenienia się TPA.
5.2.1 Tp0136
Tp0136 zidentyfikowano jako wiążący fibronektynę OMP TPA, który wiąże się z komórką skuteczniej niż fibronektyna w osoczu ( 50 , 51 ). To selektywne wiązanie obejmuje różne domeny tego białka. Zatem konserwatywny region N-końcowy jest głównie odpowiedzialny za wiązanie z fibronektyną w osoczu. Reszty na C-końcu, a także w centralnej części białka, uczestniczą w wiązaniu się z komórkową formą fibronektyny ( 51 ). Co ciekawe, Tp0136 wykazuje zmienność sekwencji pomiędzy szczepami TPA, a jego transkrypcja jest regulowana podczas infekcji ( 50 , 51 ). Interakcja Tp0136 z fibronektyną wyrażaną na powierzchni komórki może sprzyjać przyłączaniu się bakterii do tkanek, ułatwiając rozprzestrzenianie się w organizmie i sprzyjając kolonizacji dystalnych tkanek śródbłonka, centralnego układu nerwowego lub łożyska (153 ) .
Oprócz swojej funkcji adhezyjnej, Tp0136 może również odgrywać ważną rolę w gojeniu wrzodu, promując migrację fibroblastów i komórek śródbłonka mikronaczyniowego oraz aktywację i agregację płytek krwi ( 154 ).
Doświadczenia w zakresie immunizacji i prowokacji przeprowadzone na królikach przy użyciu rekombinowanego Tp0136 wytwarzanego w E. coli wykazały opóźnienie owrzodzenia zmian chorobowych, ale nie ich rozwoju, co wskazuje na brak ochrony przed infekcją ( 50 ). Co godne uwagi, surowice uzyskane od immunizowanych zwierząt zmniejszały przyleganie bakterii do fibronektyny, chociaż na niższym poziomie niż surowice uzyskane od nieszczepionych i zakażonych królików ( 50 ). Co ciekawe, Xu i in. wykazali, że króliki immunizowane Tp0136 wywołały wysoki poziom przeciwciał swoistych dla antygenu, osłabiony rozwój zmian chorobowych ze zwiększonym naciekiem komórkowym i ograniczonym rozsiewem krętków do odległych narządów ( 152 ). Chociaż istnieją rozbieżności między obydwoma badaniami, które można wyjaśnić różnicami w adiuwancie, harmonogramie immunizacji i ilości żywych zaszczepionych bakterii, wyniki te sugerują, że przeciwciała wytworzone przeciwko Tp0136 mogą działać ochronnie. Jednakże fakt, że przeciwciała powstałe podczas eksperymentalnej infekcji blokowały skuteczniej wiązanie TPA z fibronektyną niż przeciwciała wywołane szczepionką przeciwko Tp0136, sugeruje, że podczas projektowania szczepionki należy uwzględnić także inne swoistości, aby skuteczniej blokować wiązanie bakterii z macierzą pozakomórkową ( 51 ).
5.2.2 Tp0155
Opisano, że Tp0155 korzystnie wiąże się z macierzą fibronektyny i dlatego może brać udział w rozprzestrzenianiu się TPA ( 155 ). Tp0155 wykazuje dwie domeny motywów lizyny (LysM) w swojej N-końcowej części, które mogą odgrywać główną rolę w wiązaniu z fibronektyną i peptydoglikanami, oraz domenę peptydazy M23 w regionie C-końcowym, która może wykazywać aktywność lityczną peptydoglikanu (156 ) .
Istnieją pewne kontrowersje dotyczące lokalizacji powierzchni Tp0155. Testy hamowania wykazały, że rekombinowany Tp0155 zmniejsza wiązanie żywego TPA ze szkiełkami opłaszczonymi fibronektyną, co sugeruje, że Tp0155 ulega ekspresji na OM ( 155 ). Wynik ten był zgodny z faktem, że TDE2318, białko wiążące fibronektynę o wysokiej homologii z Tp0155, znaleziono na powierzchni T. denticola zgodnie z danymi z mikroskopii immunofluorescencyjnej ( 156 ). Jednakże badania immunofluorescencyjne z użyciem treponemów mikrokapsułkowanych w agarozie wykazały, że Tp0155 nie jest eksponowany na OM TPA ( 157 ). Zgodnie z tą obserwacją króliki immunizowane Tp0155, które wywołały wysokie miano przeciwciał swoistych wobec antygenu, nie były chronione przed śródskórną prowokacją żywymi krętkami ( 157 ). Zatem konieczne będą dalsze badania w celu ustalenia, czy Tp0155 będzie prawdziwym OMP, czy nie.
Co ciekawe, badania przeprowadzone na surowicy ludzkiej wykazały, że około 70% pacjentów chorych na kiłę nie wykazywało przeciwciał przeciwko Tp0155, co wskazuje, że ekspresja tego białka lub jego immunogenność może być niska w przypadku naturalnej infekcji (157, 158 ).
5.2.3 Tp0435
Tp0435 jest wysoce immunogenną lipoproteiną o masie 14 kDa z funkcją adhezyny, znaną również jako Tp17. Aby zidentyfikować lokalizację Tp0435 w TPA, Chan i wsp. przeprowadzili badanie, w którym analizowano ekspresję powierzchniową Tp0435 w TPA i komórkach B. burgdorferi transformowanych Tp0435 ( 159 ). Autorzy doszli do wniosku, że Tp0435 można modyfikować potranslacyjnie, tworząc kilka wariantów, które można w różny sposób zlokalizować pomiędzy powierzchnią a przestrzenią peryplazmatyczną. W przeciwieństwie do tego Cox i wsp. nie znaleźli dowodów na działanie powierzchniowe Tp0435 ( 160 ). Strukturalnie Tp0435 obejmuje ośmioniciową antyrównoległą beczkę β z domeną przypominającą basen umieszczoną na jednym końcu ( 161 ). Według Chan i wsp., Tp0435 sprzyja przyłączaniu się krętków do linii komórkowych ssaków ( 159 ). Co ciekawe, surowice ludzkie zakażonych pacjentów reagują przeciwko Tp0435 ( 158 ), a immunizacja królików szczepem B. burgdorferi wyrażającym Tp0435 wywołała silną odpowiedź immunologiczną przeciwko Tp0435. Jednakże ta odpowiedź immunologiczna nie chroniła przed infekcją lub rozwojem zmian chorobowych po eksperymentalnej prowokacji ( 162 ). Zatem mogą być potrzebne dalsze badania w celu określenia lokalizacji Tp0435 i jego potencjału jako kandydata na szczepionkę.
5.2.4 Tp0483
Tp0483 może wiązać zarówno macierz zewnątrzkomórkową, jak i rozpuszczalne fibronektyny ( 155 ). Część C-końcowa Tp0483 (179-374 aminokwasów) również wykazywała reaktywność wobec lamininy ( 163 ). Niewiele wiadomo na temat struktury Tp0483. Jednakże peptyd zawierający 316-333 aminokwasów hamował wiązanie Tp0483 z fibronektyną, co sugeruje, że ten peptyd lub bliski region Tp0483 powinien być odpowiedzialny za wiązanie z adhezyjną glikoproteiną (164 ) . Podobnie jak Tp0155, Cameron i in. pośrednio zasugerował, że Tp0483 był OMP ( 155 ). Przeciwnie, Tomson i współpracownicy nie znaleźli dowodów na lokalizację Tp0483 OM. Mimo że króliki immunizowane tym białkiem uzyskały wysokie miano przeciwciał, autorzy nie wykryli ochrony po prowokacji ( 157 ).
5.2.5 Tp0750
Tp0750 wiąże się z fibrynogenem i kompleksem receptora fibrynolitycznego, białkiem aneksyną A2. Tp0750 wykazuje aktywność metaloproteazy serynowej i jest zdolny do degradacji fibrynogenu i fibronektyny, hamując kaskadę krzepnięcia, ale nie może rozkładać skrzepów fibrynowych ( 57 ). Strukturalnie Tp0750 wykazuje miejsce adhezji zależnej od jonów metalu (MIDAS) zawierające domeny typu A czynnika von Willebranda (obszar V29-T147). Domeny MIDAS wiążą się z pierwotnym wapniem. Białko to, podobnie jak Tp0751 (patrz sekcja 5.2.6), podlega kotranskrypcji i może współistnieć jako kompleks heterodimeryczny. Jednakże, w przeciwieństwie do Tp0751, Tp0750 wykazuje ograniczoną zdolność wiązania lamininy ( 57 ). Analizy sekwencji obu białek wykazały, że Tp0750 jest wysoce konserwatywny wśród patogennych gatunków krętków, podczas gdy Tp0751 jest mniej konserwatywny, z wyjątkiem najbardziej inwazyjnych gatunków krętków. Co ciekawe, ortologi Tp0750 i Tp0751 z mniej inwazyjnych gatunków krętków nie wiążą się ani nie degradują białek gospodarza, co wzmacnia pogląd, że zarówno funkcja adhezji, jak i zdolność do degradacji są ważne dla progresji kiły ( 165 ).
5.2.6 Tp0751
Chociaż istnieje pewien stopień zgodności co do struktury Tp0751, istnieją rozbieżności dotyczące jego funkcji, lokalizacji i znaczenia odpowiedzi immunologicznej wywołanej przeciwko temu białku. Ze strukturalnego punktu widzenia Tp0751 przedstawia topologię dwuczęściową ze zdezorganizowanym obszarem N-końcowym, który jest połączony poprzez domenę α-helisy z częścią C-końcową ułożoną w osiem antyrównoległych arkuszy β (52 , 166 ) .
Wcześniej opisano, że Tp0751 wiąże się z różnymi formami lamininy w sposób zależny od dawki i może działać jako adhezyna naczyniowa, sprzyjając rozprzestrzenianiu się bakterii ( 53 , 167 ). Dodatkowo Tp0751 wykazywał aktywność metaloproteazy i może rozkładać fibrynogen i lamininę, sprzyjając rozpuszczaniu skrzepów i sprzyjaniu rozprzestrzenianiu się bakterii ( 58 ). Jednakże Luthra i współpracownicy wykazali, że Tp0751 znajduje się w przestrzeni peryplazmatycznej, a jego główną rolą jest wiązanie się z małymi cząsteczkami wzdłuż krawędzi beczki, takimi jak hemy (166 ) .
Podczas gdy Cameron i współpracownicy opisali, że Tp0751 jest rozpoznawany przez przeciwciała w surowicy pacjentów i zakażonych królików ( 163 ), a przeciwciała te mogą opsonizować bakterię, Luthra i in. zaobserwowali odwrotne wyniki. W ich badaniu Tp0751 indukował słabą odpowiedź przeciwciał u zakażonych ludzi i królików poddanych prowokacji, a przeciwciała anty-Tp0751 nie mają zdolności opsonizacji, prawdopodobnie z powodu niskiego poziomu ekspresji Tp0751 na OM (166 ) . Eksperymenty ze szczepieniem i prowokacją na królikach również dały sprzeczne wyniki. Lithgow i współpracownicy wykazali, że zwierzęta zaszczepione Tp0751 wykazywały osłabione zmiany chorobowe z niskim obciążeniem bakteryjnym. Chociaż szczepienie nie zapobiegło infekcji, skutecznie hamowało rozprzestrzenianie się bakterii ( 168 ). Z drugiej strony Luthra i in. nie wykazali żadnej ochrony przed miejscową lub rozsianą infekcją po śródskórnej prowokacji TPA u zwierząt immunizowanych Tp0751 ( 166 ). Dlatego też konieczne będą dalsze badania w celu wyjaśnienia tych rozbieżności.
5.2.7 Tp0954
Tp0954 został niedawno opisany jako lipoproteina powierzchniowa i może odgrywać główną rolę w kile wrodzonej ( 169 ). Zgodnie ze swoją strukturą przewiduje się, że białko to będzie miało motyw strukturalny powtórzenia tetratrikopeptydowego (TPR) z tandemowymi α-helisami. Co więcej, sekwencja Tp0954 jest zachowana w przypadku kilku szczepów TPA (np. Nichols, Chicago, Mexico A i Amoy) ( 169 ). Primus i in. opisali również, że bakteria B. burgdorferi B314 B. burgdorferi B314 transformowana Tp0954 , o której wiadomo, że jest szczepem słabo przylegającym, nawiązała wiązanie z liniami komórek nabłonkowych ssaków, w tym z linią komórek ludzkiego łożyska [tj. BeWo (CCL-98)] (169 ) . W przeciwieństwie do innych adhezyn TPA, Tp0954 może pośredniczyć w adhezji i rozprzestrzenianiu się bakterii poprzez wiązanie się z glikozaminoglikanami obecnymi w ludzkim łożysku, takimi jak siarczan dermatanu, heparyna i siarczan heparanu, które są składnikami macierzy zewnątrzkomórkowej (169 ). Siarczan dermatanu jest związany z naczyniami krwionośnymi płodu i powierzchnią syncytialną, podczas gdy siarczan heparanu jest zlokalizowany w warstwach trofoblastów ( 170 – 173 ). Zatem możliwe jest, że Tp0954 może ułatwiać wrodzone zakażenie poprzez wiązanie się z glikozaminoglikanami łożyskowymi.
5.3 Inne funkcje
Wiele białek w OM TPA nie jest powiązanych ani z funkcją transportową, ani adhezyjną.
5.3.1 Tp0326
Tp0326 (lub Tp92) wykazuje homologię z ortologami BamA innych bakterii Gram-ujemnych. Białka podobne do BamA charakteryzują się dwuczęściową strukturą z domeną β-beczułki zlokalizowaną w części C-końcowej i co najmniej jedną domeną związaną z transportem polipeptydu (POTRA) na końcu N. W szczególności przewidywano, że Tp0326 będzie zawierał pięć N-końcowych domen POTRA i jedną C-końcową domenę β-baryłkową ( 174 , 175 ). Według Desrosiers i wsp. N-końcowe domeny POTRA są periplazmatyczne, podczas gdy C-końcowa beczka β jest osadzona w OM tworząc por (175). N -końcowy koniec może wiązać się z wieloma białkami periplazmatycznymi poprzez domeny POTRA , tworząc kompleks białkowy kluczowy dla syntezy błony. Sugeruje się, że podobnie jak inne białka podobne do BamA, Tp0326 jest częścią maszynerii białkowej zaangażowanej w translokację i insercję białek z przestrzeni peryplazmatycznej do TPA OM (175 ) . Co ciekawe, przeciwciała ludzkie są skierowane głównie przeciwko segmentowi peryplazmatycznemu Tp0326, podczas gdy przeciwciała królicze celują zarówno w regiony N-końcowe, jak i C-końcowe ( 175 ). Rzeczywiście, Luthra i in. odkryli, że domeny POTRA immunodominują nad domenami β-beczułki zarówno u królików, jak i u ludzi ( 174 ). Ponadto autorzy ci opisali, że centralny por Tp0326 odsłania osiem pętli do przestrzeni zewnątrzkomórkowej. Wśród nich pętla 4 jest szczególnie celem przeciwciał o zdolnościach opsonizujących pochodzących od zakażonych królików i ludzi chorych na kiłę wtórną ( 174 ). Natomiast Tomsonowi i college’om nie udało się wykryć ekspresji Tp0326 na OM krętków zamkniętych w mikrokropelkach żelu agarozowego przy użyciu pośredniej immunofluorescencji ( 157 ). Ponadto nie zaobserwowali ochrony u zwierząt immunizowanych Tp0326 po prowokacji TPA. Z drugiej strony Cameron i in. wykazali, że chociaż wszystkie króliki immunizowane Tp0326 zostały zakażone po prowokacji, u zaszczepionych zwierząt, które miały wysokie miano przeciwciał swoistych wobec antygenu, występował pewien stopień ochrony ( 176 ).
5.3.2 Tp0257
Tp0257 (lub GpD) zidentyfikowano przy użyciu biblioteki ekspresji genetycznej krętków i opsonizowanej surowicy odpornościowej królika. Białko to wykazuje homologię z białkiem GplQ H. influenzae , białkiem fosfodiesterazy glicerolowo-fosfodiestrowej, które ulega częściowej ekspresji na powierzchni ( 177 ). Zatem możliwe jest, że Tp0257 może być białkiem OM. W związku z tym Cameron i in. zgłosili pośrednie dowody jego lokalizacji na powierzchni za pomocą analizy immunoblot lizatów TPA. Autorzy zaobserwowali pozytywne barwienie w preparacie zawierającym OM, ale nie we frakcji lizatu TPA usuniętej z OM ( 178 ). Co więcej, z preparatów OM wyizolowano GpD [ 179 ]. Co ciekawe, króliki immunizowane rekombinowanym Tp0257 wykazywały zmniejszenie rozwoju zmian chorobowych i proliferacji bakterii po prowokacji TPA ( 178 ). Z drugiej strony Shevchenko i wsp. wykazali, że Tp0257 może być białkiem peryplazmatycznym, ponieważ w wielu testach nie zidentyfikowali dowodów eksponowanych na powierzchni, a immunizacja Tp0257 nie chroniła królików przed prowokacją TPA (180 ) . Niedawno szczepionka DNA kodująca białko fuzyjne Tp0257-IL-2 wykazała zmniejszenie zmian wrzodziejących, a także liczby zmian zawierających TPA ( 181 ). Włączenie IL-2 do szczepionki zwiększyło odpowiedź humoralną anty-GpD i poziom IFN-γ. Co godne uwagi, króliki immunizowane szczepionką pierwotną DNA Tp0257-IL2 i donosową dawką przypominającą rekombinowanym białkiem z adiuwantem indukowanym CpG-ODN odpornością śluzówkową i ogólnoustrojową i wykazywały szybszy powrót do zdrowia po wszczepieniu TPA (182 ) .
Co ciekawe, Tp0257 jest konserwatywny wśród szczepów TPA, jak również u Treponema pallidum endemicum i Treponema pallidum pertenue ( 183 ). Wyniki te czynią to białko atrakcyjnym kandydatem na immunogen do projektowania szczepionek.
5.3.3 Tp1038
Tp1038, znany również jako TpF1 lub antygen 4D, jest bakterioferrytyną o aktywności ferroksydazy, odgrywającą główną rolę w wychwycie żelaza ( 184 ). Radolf i in. wykazali, że Tp1038 jest eksponowany na powierzchni, ponieważ surowice królików immunizowanych Tp1038 reagowały na unieruchomiony TPA w obecności dopełniacza ( 185 ).
Tp1038 jest zdolny do wywoływania różnorodnych odpowiedzi immunologicznych. Na przykład białko to może brać udział w odpowiedzi prozapalnej podczas kiły pierwotnej, ponieważ może aktywować kompleks inflamasomowy w monocytach i w ten sposób indukować uwalnianie IL-1β, IL-6 i TNF-α ( 186 , 187 ) . , co może skutkować uszkodzeniem tkanek związanym z tym etapem. Jednakże Babolin i in. opisali, że Tp1038 może również powodować odpowiedź regulacyjną T ( 186 ). Komórki T CD4 + CD25 + Foxp3 + od pacjentów z kiłą wtórną wytwarzają TGF-β pod stymulacją Tp1038, a monocyty stymulowane Tp1038 wytwarzają IL-10 i TGF-β, dwie cytokiny powiązane z różnicowaniem komórek Treg ( 186 ) . Zatem TpF1 może być czynnikiem zjadliwości związanym z utrzymywaniem się zakażenia TPA poprzez zmniejszenie odpowiedzi immunologicznej ( 186 ). Jeśli chodzi o odpowiedź humoralną, przeciwciała anty-Tp1038 są wykrywalne we wszystkich stadiach kiły ( 186 , 188 ). Wreszcie Pozzobon i wsp. odkryli, że pacjenci z kiłą trzeciorzędową mają limfocyty T specyficzne dla Tp1038, które stymulują monocyty i komórki śródbłonka ludzkiej żyły pępowinowej do wytwarzania czynnika tkankowego, IL-8 i CCL-20 (189 ) . Cytokiny te biorą udział w angiogenezie, zatem Tp1038 może mieć także związek z kiłą trzeciorzędową, w której zmiany chorobowe na tym etapie charakteryzują się zapaleniem naczyń i angiogenezą ( 189 ). Zatem Tp1038 jest interesującym celem w opracowywaniu szczepionek, ponieważ bierze udział we wszystkich stadiach kiły.
6 Strategie unikania odporności
Chociaż odpowiedź immunologiczna jest wyzwalana przeciwko TPA, nie chroni ona przed rozprzestrzenianiem się bakterii i postępem choroby do drugiego lub trzeciego stadium. Ponadto reinfekcje są dość częste i występują z częstością około 5–22% ( 190–192 ) , co wskazuje, że TPA rozwinęło pewne mechanizmy pozwalające uniknąć układu odpornościowego ( ryc. 2 ).
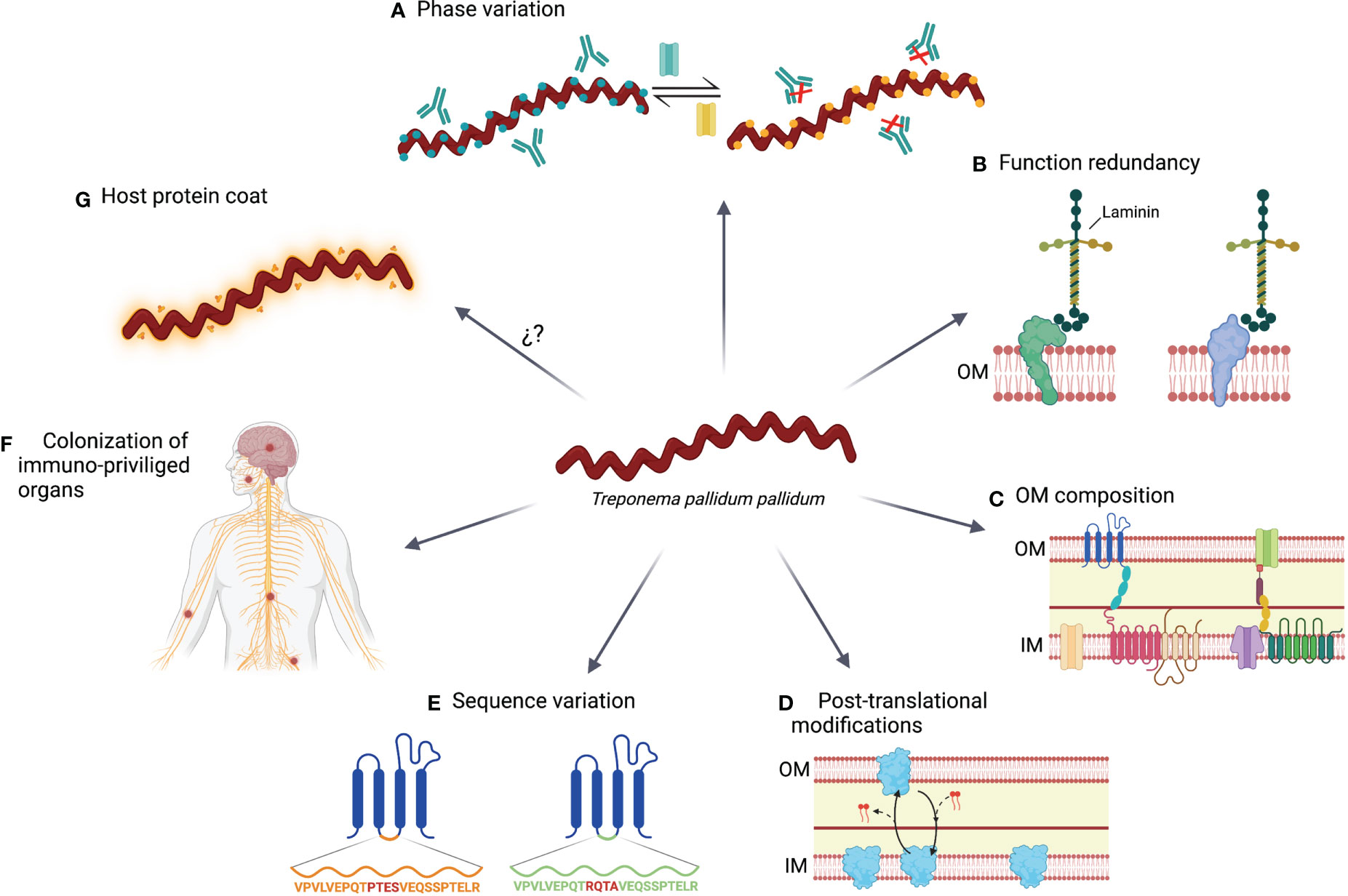
6.1 Zmiana fazy lub włączanie/wyłączanie
TPA może modulować ekspresję kilku OMP podczas swojego cyklu życia ( 122 ). Jednakże mechanizmy kontrolujące ekspresję tych białek nie są dobrze scharakteryzowane. W przypadku podrodzin I i II białek Tpr, powyżej miejsca startu transkrypcji zidentyfikowano powtórzenia homopolimeru guanozyny (poliG). Długość sekwencji poliG różni się w przypadku izolatów krętkowych i moduluje ekspresję białek Tpr na poziomie transkrypcji ( 124 ). Sygnały PolyG znaleziono także w innych OMP, takich jak białko Tp0126 ( 140 ). Zmienność fazową opartą na hiperzmiennych powtórzeniach homopolimerowych opisano wcześniej u kilku innych bakterii, takich jak Neisseria meningitidis , Helicobacter pylori lub Haemophilus influenzae , i ma ona kluczowe znaczenie dla przeżycia patogenów stojących w obliczu odporności gospodarza ( 193 – 195 ). Ponadto zidentyfikowano inne mechanizmy regulacyjne transkrypcji, takie jak elementy DNA działające w układzie cis G4FS, które tworzą kwadrupleksy guaniny i indukują rekombinację i konwersję genu Tp0136 ( 196 ). Korzystając z tych mechanizmów, TPA może modyfikować swój repertuar OMP, aby dostosować się do warunków środowiskowych i uniknąć odpowiedzi immunologicznej. W rezultacie specyficzność odpowiedzi humoralnej przeciwko białkom Tpr może różnić się u zwierząt i ludzi zakażonych tym samym lub innym szczepem TPA, co potwierdza istnienie zróżnicowanego repertuaru OMP (81 ) .
6.2 Redundancja funkcjonalna
Kilka OMP pełni podobne funkcje w transporcie lub adhezji komórek. Redundancja funkcjonalna może być korzystna, ponieważ zastąpienie jednego białka funkcjonalnym homologiem może zagwarantować, że zmienność fazowa i modyfikacje potranslacyjne nie wpłyną niekorzystnie na homeostazę komórki ( 22 ). Jako przykład doniesiono , że Salmonella ma dwa różne geny flageliny, fliC i fliB, o tej samej funkcji, ale innej strukturze. Zmieniając ekspresję jednego białka flageliny na drugie, układ odpornościowy staje się nieskuteczny wobec złożonej wici ( 197 ).
6.3 Skład błony zewnętrznej
TPA charakteryzuje się małą liczbą OMP, co może wiązać się z zaletą w postaci ograniczenia liczby antygenów eksponowanych na układ odpornościowy. Co więcej, rozmieszczenie tych białek może sprzyjać temu, że przeciwciała wiążą się z nimi tylko jednym ramieniem, zmniejszając zachłanność interakcji. Mechanizm ten opisano wcześniej w zakażeniu ludzkim wirusem niedoboru odporności (HIV) ( 198 ), które również charakteryzuje się niską gęstością glikoproteiny otoczki na powierzchni wirionu. W przypadku HIV polireaktywność przeciwciał neutralizujących może zwiększać awidność wiązania poprzez heteroligację ze składnikami błony ( 198 ). Jednakże rozwój tych przeciwciał może być ograniczony przez mechanizmy tolerancji ( 199 ). To, czy zjawisko to jest również powiązane z odpornością na TPA, wymaga dalszych badań, ale może częściowo wyjaśniać niską skuteczność przeciwciał w kontrolowaniu infekcji. Ponieważ OM nie zawiera LPS ( 27 , 28 ) i jest wzbogacona w fosfolipidy o działaniu przeciwzapalnym (np. fosfatydyloserynę i fosfatydylocholinę), cechy te mogą również utrudniać rozwój skutecznej odpowiedzi immunologicznej ( 200 , 201 ).
6.4 Modyfikacje potranslacyjne
Potranslacyjne modyfikacje białek odgrywają kluczową rolę w różnych procesach komórek prokariotycznych, w tym w trwałości i zjadliwości ( 202 ). Modyfikacje te wykryto w proteomie TPA. Na przykład istnieje kilka wariantów Tp0435, które mogą współistnieć i zostały wytworzone w wyniku palmitoilacji ( 159 ). Palmitoilacja polega na dodaniu łańcuchów lipidowych do końcowych reszt cysteiny. Proces ten opisano wcześniej u innych bakterii i przyczynia się on do ich zakaźności i rozpoznawania przez układ odpornościowy ( 203 ). Warianty Tp0435 mogą być zlokalizowane na powierzchni lub w przestrzeniach peryplazmatycznych, przy czym ta ostatnia jest najliczniejsza ( 159 ), co może wyjaśniać brak konsensusu co do lokalizacji Tp0435. W rzeczywistości białko to było pierwszym opisem zmodyfikowanych potranslacyjnie wariantów białka w TPA. Jednak inne OMP mogą być również podatne na modyfikacje potranslacyjne.
6.5 Zmiana sekwencji
W TPA opisano genetyczną zmienność kilku OMP, w tym szczepów przystosowanych laboratoryjnie lub pierwotnie izolowanych ( 127 , 129 , 204 ). Niektóre z tych zmian opisano w rodzinie Tpr, obejmującej białka TprC, TprD, TprG i TprK ( 127 , 204 , 205 ). Co ciekawe, TprK gromadzi zmienność sekwencji w regionach zmiennych (szczególnie w V6) poprzez niewzajemną rekombinację z cichymi kasetami, która jest wzmacniana pod presją immunologiczną ( 129 , 130 ). Dodatkowo zidentyfikowano kilka alleli tprk w różnych szczepach TPA ( 127 ), co przyczynia się do zwiększenia zmienności tego białka. Oprócz białek Tpr inne OMP, takie jak Tp0326 (BamA), również wykazują warianty genetyczne ( 204 , 205 ). W tym białku nagromadziły się mutacje w zewnątrzkomórkowych pętlach hydrofilowych, które, jak się przewiduje, zawierają epitopy dla komórek B ( 204 ). Zmienność sekwencji potwierdzono także w białkach ortologów FadL w zakresie od w pełni konserwatywnego Tp0856 do głęboko zmiennego Tp0548, jak również w Tp0136, Tp0868, Tp0966 i Tp0967 ( 206 – 208 ).
6.6 Kolonizacja narządów uprzywilejowanych pod względem odporności
TPA może kolonizować dalsze narządy o uprzywilejowanej odporności, takie jak centralny układ nerwowy, łożysko lub oczy. W tych tkankach działanie układu odpornościowego może być ograniczone, przyczyniając się do utrzymywania się tego patogenu ( 209 – 211 ). W rzeczywistości badania przeprowadzone na modelu królika wykazały, że TPA można wykryć w nerwach wcześnie po zakażeniu ( 68 ).
6.7 Płaszcz białek gospodarza
Podobnie jak inne bakterie; na przykład u niektórych gatunków paciorkowców ( 212 ) unikanie odporności TPA może być związane z białkami gospodarza tworzącymi powłokę powierzchniową, znaną również jako przebranie antygenowe. Alderete i Baseman opisali, że zarówno albuminy królicze, jak i ludzkie, a także inne białka gospodarza, mogą być adsorbowane na powierzchni TPA ( 213 ). Egesten i in. wykazali, że paciorkowce grupy G wykazują ekspresję białka G na swojej powierzchni i jest ono zdolne do wiązania ludzkiej albuminy, a zatem hamuje przeciwdrobnoustrojowe działanie CXCL9 ( 214 ). W związku z tym zaproponowano, że zdolność Tp0483 do wiązania rozpuszczalnej fibronektyny może być powiązana z unikaniem odpowiedzi humoralnej ( 164 ). Jednakże Cox i wsp. nie wykryli białek surowicy na powierzchni TPA za pomocą mikroskopii immunoelektronowej ( 26 ), stwierdzając, że niska gęstość OMP może być głównym mechanizmem unikania odporności TPA.
7 Dyskusja
Kiła jest wielostopniową chorobą wywoływaną przez TPA, na którą nie ma skutecznej szczepionki. Pomimo kilkudziesięciu lat badań, niemożność genetycznej manipulacji i hodowli TPA in vitro , niedostatek OMP, kruchość OM i brak odpowiedniego modelu zwierzęcego, który podsumowuje wszystkie stadia ludzkiej kiły, utrudniają postęp w tej dziedzinie. Ponadto wiele kontrowersji pozostaje nierozwiązanych w odniesieniu do identyfikacji OMP, lokalizacji, funkcji, potencjału tych białek do indukowania ochronnych odpowiedzi immunologicznych lub charakterystyki tych odpowiedzi. Naukowcy postawili hipotezę, że odpowiedź immunologiczna na DTH jest niezbędna do kontrolowania i usuwania infekcji TPA, ale jest również odpowiedzialna za rozwój uszkodzeń tkanek, w tym wrzodziejących zmian tworzących wrzody. Ponieważ w DTH pośredniczą głównie limfocyty T Th1 CD4 + , kluczowa jest prezentacja antygenów przez DC, makrofagi lub inne profesjonalne komórki prezentujące antygen. Warto zauważyć, że proces ten jest niezależny od lokalizacji antygenu i zarówno antygeny wewnątrzkomórkowe, jak i eksponowane na powierzchni mogą być niewyraźnie przetwarzane i prezentowane limfocytom T. Dlatego oba antygeny mogą generować ochronne odpowiedzi limfocytów T Th1 CD4 + . Zatem u królików immunizowanych wewnątrzkomórkowymi antygenami endoflagellarnymi, które zostały bezpośrednio wyizolowane z TPA, wykazano szybszy rozwój zmian chorobowych ( 215 ). Jednakże w zmianach tych nie wykryto bakterii, co można wytłumaczyć przyspieszoną odpowiedzią DTH w pamięci. Natomiast króliki immunizowane plazmidem kodującym FlaB3 wykazywały osłabiony rozwój zmian chorobowych, który był związany ze zwiększonym naciekiem komórkowym i hamowaniem rozprzestrzeniania się bakterii do narządów dalszych (216 ) . Zgodnie z tą obserwacją króliki immunizowane TprF, białkiem peryplazmatycznym, wykazywały osłabione zmiany skórne i zmniejszone obciążenie bakteryjne ( 136 ). Przyczyny tych różnych wyników nie są w pełni zrozumiałe, ale mogą mieć to związek z różnicami w składzie szczepionki i drogach podawania, wielkości odpowiedzi komórek T indukowanych szczepionką oraz względnej reprezentacji antygenów (flagelliny i TprF) w całym proteomie TPA. Podobne wyniki do immunizacji TprF zaobserwowano również, gdy jako antygeny zastosowano OMP. Dlatego DTH może być ochronną odpowiedzią immunologiczną, która modyfikuje rozwój zmian chorobowych i usuwa bakterie z miejsc infekcji. Jednakże żadne z białek TPA testowanych dotychczas jako immunogeny nie chroniło przed infekcją, co wskazuje, że prawdopodobnie będą wymagane dodatkowe odpowiedzi immunologiczne.
Pomimo pewnych kontrowersji [ 68 ], mocne dowody potwierdzają ochronną rolę przeciwciał, ułatwiając opsonizację i zabijanie bakterii przez wrodzone komórki odpornościowe, a także blokując ich interakcję ze składnikami macierzy zewnątrzkomórkowej, takimi jak laminina i fibronektyna. Funkcje te ściśle zależą od ekspozycji antygenów na OM oraz, w niektórych przypadkach (np. fagocytoza bakteryjna), od interakcji z receptorami Fcγ wyrażanymi na powierzchni komórek odporności wrodzonej, takich jak makrofagi. Interakcja IgG-FcγR może także polepszyć prezentację antygenu przez komórki prezentujące antygen i generowanie odpowiedzi komórkowych Th1 ( 217 ). Dlatego też, aby opracować ochronną szczepionkę TPA, kluczowe jest określenie, które typy antygenów mogą indukować synergiczną odpowiedź immunologiczną skierowaną przeciwko TPA. Istotne jest również rozważenie strategii unikania odporności opracowanych przez TPA, takich jak redundancja funkcjonalna i zmienność fazowa. W tym sensie nakierowanie na kilka białek spełniających dwie lub więcej funkcji, takich jak transport i adhezja, mogłoby zwiększyć skuteczność szczepionki, jak zaobserwowano wcześniej ( 218 , 219 ).
Idealna szczepionka przeciwko kile powinna zapewniać ochronę przed zakażeniem TPA. Jednakże nawet jeśli szczepionka nie jest w stanie całkowicie zapobiec infekcji, nadal warto ją rozważyć jako środek ograniczający rozprzestrzenianie się bakterii i inwazyjność tkanek, blokujący powstawanie latencji lub zapobiegający postępowi choroby do drugiego i trzeciorzędowego stadium (113 ) . Ponadto szczepionka ograniczająca trwałość bakterii lub zapobiegająca inwazji TPA przez łożysko może również zmniejszyć liczbę przypadków kiły wrodzonej na obszarach endemicznych, gdzie liczba niezdiagnozowanych osób zakażonych może być wysoka, a dostęp do opieki zdrowotnej może być ograniczony. Co więcej, szczepionka przeciw kile musi być skuteczna przeciwko możliwym ponownym zakażeniom podobnymi lub różnymi szczepami lub izolatami TPA, co wskazuje na znaczenie stosowania konserwatywnych antygenów. Ostatnio pojawiły się dowody na to, że TPA można hodować in vitro i że można dokonać manipulacji genetycznej tego patogenu ( 114 , 220 , 221 ). Te postępy naukowe prawdopodobnie otworzą drogę do nowych kierunków badań, które mogą rzucić światło na obecne kontrowersje.
Autorskie Wkłady
Treść naukową zaprojektowali CÁ-N, OM, BC, JB i JC. CÁ-N, NP-L i JC zredagowali rękopis i ryciny. OM, BC i JB zredagowali manuskrypt i uczestniczyli w dyskusji merytorycznej. Wszyscy autorzy przeczytali i zaakceptowali treść manuskryptu.
Finansowanie
Prace te były wspierane przez Program CERCA (2017 SGR 252; Generalitat de Catalunya), Direcció General de Recerca i Innovació en Salut (Generalitat de Catalunya) (projekty SLD0015 i SLD0016), Instytut Zdrowia Carlosa III (PI17/01518 i PI18/ 01332). JB jest wspierany przez Departament Zdrowia Rządu Katalonii (Generalitat de Catalunya). CÁ-N otrzymał stypendium przeddoktorskie od Generalitat de Catalunya i Fons Social Europeu (2020 FI_B_0742). Badanie to było również wspierane przez CIBER – Consorcio Centro de Investigación Biomédica en Red (CB 2021), Instytut Zdrowia Carlosa III, Ministerio de Ciencia e Innovación i Unión Europea – NextGenerationEU. Fundatorzy nie mieli żadnej roli w projektowaniu badania, analizie danych i podejmowaniu decyzji o publikacji lub przygotowanie manuskryptu.
Podziękowanie
Rysunek 1 i Rysunek 2 zostały utworzone za pomocą BioRender.com.
Konflikt interesów
Autorzy oświadczają, że badanie przeprowadzono w warunkach braku powiązań handlowych lub finansowych, które mogłyby stanowić potencjalny konflikt interesów.
Notatka wydawcy
Wszystkie roszczenia wyrażone w tym artykule są wyłącznie roszczeniami autorów i niekoniecznie reprezentują roszczenia organizacji z nimi stowarzyszonych lub wydawcy, redaktorów i recenzentów. Żaden produkt, który może być oceniany w tym artykule, lub twierdzenie, które może być wysunięte przez jego producenta, nie jest objęte gwarancją ani poparciem wydawcy.
Bibliografia
1. Harper KN, Zuckerman MK, Harper ML, Kingston JD, Armelagos GJ. Ponowne spojrzenie na pochodzenie i starożytność kiły: ocena prekolumbijskich dowodów starego świata na zakażenie krętkiem. Am J Phys Anthropol (2011) 146: 99–133. doi: 10.1002/ajpa.21613
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
2. De Melo FL, De Mello JCM, Fraga AM, Nunes K, Eggers S. Syphilis na skrzyżowaniu filogenetyki i paleopatologii. PloS Negl Trop Dis (2010) 4:e575. doi: 10.1371/journal.pntd.0000575
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
3. Mahoney JF, Arnold RC, Harris A. Leczenie kiły wczesnej penicyliną – doniesienie wstępne. Am J. Public Health Nations Health (1943) 33:1387–91. doi: 10.2105/ajph.33.12.1387
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
4. Singh AE, Romanowski B. Syphilis: Przegląd ze szczególnym uwzględnieniem cech klinicznych, epidemiologicznych i niektórych biologicznych. Recenzje mikrobiologii klinicznej (1999) 12: 187–209. doi: 10.1128/cmr.12.2.187
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
5. Peeling RW, Mabey D, Kamb ML, Chen XS, Radolf JD, Benzaken AS. Syfilis. Nature Recenzje Podstawy chorób (2017) 3:17073. doi: 10.1038/nrdp.2017.73
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
6. Radolf JD, Deka RK, Anand A, Šmajs D, Norgard MV, Yang XF. Treponema pallidum, krętek kiły: zarabianie na życie jako ukryty patogen. Nat Rev Microbiol (2016) 14:744–59. doi: 10.1038/nrmicro.2016.141
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
7. Denee Thomas D, Navabo M, Haaket DA, Fogelmant AM, Miller JN, Lovett MA. Treponema pallidum atakuje połączenia międzykomórkowe monowarstw komórek śródbłonka. Proc Natl Acad Sci USA (1988) 85(10):3608–12. doi: 10.1073/pnas.85.10.3608
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
8. Lafond RE, Lukehart SA. Biologiczne podłoże kiły. Clin Microbiol Rev (2006) 19:29–49. doi: 10.1128/cmr.19.1.29-49.2006
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
9. Cruz AR, Ramirez LG, Zuluaga AV, Pillay A, Abreu C, Valencia CA i in. Unikanie odporności i rozpoznawanie krętka kiły we krwi i skórze pacjentów z kiłą wtórną: dwa immunologicznie odrębne przedziały. PloS Negl Trop Dis (2012) 6(7):e1717. doi: 10.1371/journal.pntd.0001717
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
10. Zaczep EW. Syfilis. Lancet (2017) 389:1550–7. doi: 10.1016/S0140-6736(16)32411-4
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
11. Luo Z, Zhu L, Ding Y, Yuan J, Li W, Wu Q i in. Czynniki związane z niepowodzeniem leczenia kiły i ponownym zakażeniem: podłużne badanie kohortowe w Shenzhen w Chinach. BMC Infect Dis (2017) 17: 1–5. doi: 10.1186/s12879-017-2715-z
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
12. Brewer TH, Peterman TA, Newman DR, Schmitt K. Reinfekcje podczas epidemii kiły na Florydzie, 2000-2008. Sex Transm Dis (2011) 38:12–7. doi: 10.1097/OLQ.0b013e3181e9afc7
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
13. Fakty dotyczące chorób przenoszonych drogą płciową – kiła (szczegółowo). Dostępne pod adresem: https://www.cdc.gov/std/syphilis/stdfact-syphilis-detailed.htm (dostęp: 18 marca 2020 r.).
14. Rowley J, Vander Hoorn S, Korenromp E, Low N, Unemo M, Abu-Raddad LJ i in. Chlamydia, rzeżączka, rzęsistkowica i kiła: globalne szacunki dotyczące częstości występowania i zapadalności, 2016. Bull World Health Organ (2019) 97(8):548–62. doi: 10.2471/blt.18.228486
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
15. Korenromp Id EL, Rowley J, Alonso M, Mello MB, Saman N, Id W i in. Globalne obciążenie kiłą matczyną i wrodzoną oraz powiązane niekorzystne wyniki porodów – szacunki na rok 2016 i postęp od 2012 r. PloS One (2019) 14(2):e0211720. doi: 10.1371/journal.pone.0211720
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
16. Lawn JE, Blencowe H, Waiswa P, Amouzou A, Mathers C, Hogan D i in. Poronienia martwe: wskaźniki, czynniki ryzyka i przyspieszenie do roku 2030. Lancet (2016) 387:587–603. doi: 10.1016/S0140-6736(15)00837-5
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
17. Kojima N, Klausner JD. Aktualizacja światowej epidemiologii kiły. Curr Epidemiol Rep (2018) 5(1):24–38. doi: 10.1007/s40471-018-0138-z
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
18. Ghanem KG, Ram S, Rice PA. Współczesna epidemia kiły. New Engl J Med (2020) 382: 845–54. doi: 10.1056/nejmra1901593
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
19. Europejskie Centrum Zapobiegania i Kontroli Chorób. Kiła i kiła wrodzona w Europie – przegląd trendów epidemiologicznych (2007–2018) i możliwości reagowania . Sztokholm: ECDC. (2019). doi: 10.2900/578824
20. Izard J, Renken C, Hsieh CE, Desrosiers DC, Dunham-Ems S, La Vake C i in. Tomografia krioelektronowa wyjaśnia strukturę molekularną treponema pallidum, krętka kiły. J Bacteriol (2009) 191: 7566–80. doi: 10.1128/jb.01031-09
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
21. Madigan MT, Martinko JM, Parker J. Brock Biologia mikroorganizmów . Wydanie 8. Nowy Jork: Prentice Hall International, Inc. (1997).
22. Radolf JD, Kumar S. Błona zewnętrzna treponema pallidum. Curr Top Microbiol Immunol (2018) 415: 1–38. doi: 10.1007/82_2017_44
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
23. Radolf JD, Robinson EJ, Bourell KW, Akins DR, Porcella SF, Weigel LM i in. Charakterystyka błon zewnętrznych wyizolowanych z treponema pallidum, krętka kiły. Infect Immun (1995) 63:4244–52. doi: 10.1128/iai.63.11.4244-4252.1995
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
24. Bailey DMJ. Zewnętrzna błona treponema pallidum: znaczenie biologiczne i właściwości biochemiczne. J Gen Microbiol (1985) 131(9):2349–57. doi: 10.1099/00221287-131-9-2349
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
25. Radolf JD, Norgardt MV, Schulz WW. Ultrastruktura błony zewnętrznej wyjaśnia ograniczoną antygenowość zjadliwego krętka bladego. Proc Natl Acad Sci USA (1989) 86(6):2051–5. doi: 10.1073/pnas.86.6.2051
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
26. Cox DL, Chang P., McDowall AW, Radolf JD. Błona zewnętrzna, a nie otoczka białek gospodarza, ogranicza antygenowość zjadliwego krętka bladego. Infect Immun (1992) 60:1076–83. doi: 10.1128/iai.60.3.1076-1083.1992
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
27. Hardy PH, Levin J. Brak endotoksyny u borrelia hispanica i treponema pallidum1. Proc Soc Exp Biol Med (1983) 174(1):47–52. doi: 10.3181/00379727-174-41702
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
28. Fraser CM, Norris SJ, Weinstock GM, White O, Sutton GG, Dodson R i in. Pełna sekwencja genomu Treponema pallidum, krętka kiły. Nauka (1998) 281: 375–88. doi: 10.1126/science.281.5375.375
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
29. Blanco DR, Miller JN, Lovett MA. Antygeny powierzchniowe krętka kiły i ich potencjał jako determinanty zjadliwości. Emerg Infect Dis (1997) 3: 11–20. doi: 10.3201/eid0301.970102
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
30. Charon NW, Cockburn A, Li C, Liu J, Miller KA, Miller MR i in. Unikalny paradygmat ruchliwości i chemotaksji krętków. Annu Rev Microbiol (2012) 66:349–70. doi: 10.1146/ANNUREV-MICRO-092611-150145
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
31. Liu J, Howell JK, Bradley SD, Zheng Y, Hong Zhou Z, Norris SJ. Architektura komórkowa treponema pallidum: nowa wici, stożek peryplazmatyczny i otoczka komórkowa, jak ujawniono za pomocą tomografii krioelektronowej. J. Mol Biol (2010) 403(4):546–61. doi: 10.1016/j.jmb.2010.09.020
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
32. Salazar JC, Rathi A, Michael NL, Radolf JD, Jagodziński LL. Ocena kinetyki rozprzestrzeniania się treponema pallidum do krwi i tkanek w kile eksperymentalnej metodą ilościowej reakcji PCR w czasie rzeczywistym. Infect Immun (2007) 75:2954–8. doi: 10.1128/iai.00090-07
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
33. Harman M, Vig DK, Radolf JD, Wolgemuth CW. Lepka dynamika boreliozy i krętków kiły ujawnia moment obrotowy i opór wici. Biophys J. (2013) 105:2273–80. doi: 10.1016/J.bpj.2013.10.004
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
34. Edmondson DG, Hu B, Norris SJ. Długoterminowa hodowla in vitro krętka kiły treponema pallidum subsp. blady. mBio (2018) 9(3):e01153–18. doi: 10.1128/mbio.01153-18
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
35. Lukehart SA, Marra CM. Izolacja i utrzymanie laboratoryjne treponema pallidum. Curr Protoc Microbiol (2007) 7: 12A.1.1–12A.1.18. doi: 10.1002/9780471729259.mc12a01s7
36. Willcox RR, Guthe T. Treponema pallidum. przegląd bibliograficzny morfologii, kultury i przetrwania t. pallidum i organizmy z nimi powiązane. Bull Światowy Organ Zdrowia (1966) 35(1):1–16.
37. Izzat NN, Knox JM, Werth JA, Dacres WG. Ewolucja wrzodów syfilitycznych ze zjadliwym treponema pallidum u królika. Brit J. Vener Dis (1970) 47:67. doi: 10.1136/sti.47.2.67
38. Baker-Zander S, Sell S. Badanie histopatologiczne i immunologiczne przebiegu kiły u doświadczalnie zakażonego królika. wykazanie długotrwałej odporności komórkowej. Am J. Pathol (1980) 101: 387–413.
39. Hanff PA, Fehniger TE, Miller JN, Lovett MA. Humoralna odpowiedź immunologiczna u ludzi na kiłę na polipeptydy Treponema pallidum. J. Immunol (1982) 129(3):1287–91.
40. Esteves PJ, Abrantes J, Baldauf HM, BenMohamed L, Chen Y, Christensen N i in. Szerokie zastosowanie królików jako modeli chorób człowieka. Exp Mol Med (2018) 50(5):1–10. doi: 10.1038/s12276-018-0094-1
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
41. Tantalo LC, Lukehart SA, Marra CM. Różnice specyficzne dla szczepu Treponema pallidum w neuroinwazji i fenotypie klinicznym w modelu królika. J. Infect Dis (2005) 191(1):75–80. doi: 10.1086/426510
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
42. Tansey C, Zhao C, Hopkins A, Ritter JM, Fakile YF, Pillay A i in. Model kiły przenoszonej przez odbytnicę u naczelnych innych niż człowiek. J. Infect Dis (2018) 217(7):1139–44. doi: 10.1093/infdis/jix669
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
43. Lu S, Zheng K, Wang J, Xu M, Xie Y, Yuan S i in. Charakterystyka rozprzestrzeniania się treponema pallidum u myszy C57BL/6. Przód Immunol (2021) 11:577129/bibtex. doi: 10.3389/fimmu.2020.577129/bibtex
44. Čejková DC, Zobaníková M, Chen L, Pospíšilová P, Strouhal M, Qin X i in. Sekwencje całego genomu trzech Treponema pallidum ssp. szczepy pertenue: treponemy Yaws i syphilis różnią się mniej niż 0,2% sekwencji genomu. PloS Negl Trop Dis (2012) 6(1):e1471. doi: 10.1371/journal.pntd.0001471
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
45. Mikalová L, Strouhal M, Čejková DC, Zobaníková M, Pospíšilová P, Norris SJ i in. Analiza genomu Treponema pallidum subsp. pallidum i subsp. szczepy pertenue: Większość różnic genetycznych jest zlokalizowana w sześciu regionach. PloS One (2010) 5:15713. doi: 10.1371/journal.pone.0015713
47. Ho EL, Lukehart SA. Kiła: wykorzystanie nowoczesnych podejść do zrozumienia starej choroby. J. Clin Invest (2011) 121:4584. doi: 10.1172/jci57173
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
48. Magnuson HJ, Eagle H, Fleischman R. Minimalne zakaźne inokulum krętka pallida (szczep Nichols) i rozważenie szybkości jego namnażania in vivo . Am J. Syph Gonorrhea Vener Dis (1948) 32(1):1–18.
49. Cumberland MC, Turner TB. Szybkość namnażania Treponema pallidum u królików zdrowych i odpornych. Am J. Syph Gonorrhea Vener Dis (1949) 33(3):201–12.
50. Brinkman MB, McGill MA, Pettersson J, Rogers A, Matějková P, Šmajs D i in. Nowy antygen Treponema pallidum, TP0136, jest białkiem błony zewnętrznej, które wiąże ludzką fibronektynę. Infect Immun (2008) 76(5):1848–57. doi: 10.1128/iai.01424-07
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
51. Ke W, Molini BJ, Lukehart SA, Giacani L. Treponema pallidum subsp. pallidum TP0136 jest heterogenne wśród izolatów i wiąże fibronektynę komórkową i osoczową poprzez swój koniec NH2. PLOS Zaniedbane choroby tropikalne (2015) 9(3):. doi: 10.1371/journal.pntd.0003662
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
52. Parker ML, Houston S, Pětrošová H, Lithgow KV, Hof R, Wetherell C i in. Struktura treponema pallidum Tp0751 (Pallilysin) ujawnia niekanoniczny fałd lipokalinowy, który pośredniczy w adhezji do składników macierzy zewnątrzkomórkowej i interakcjach z komórkami gospodarza. PloS Pathog (2016) 12(9):e1005919. doi: 10.1371/journal.ppat.1005919
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
53. Cameron CE, Brouwer NL, Tisch LM, Kuroiwa JMY. Określenie interakcji adhezyny treponema pallidum Tp0751 z lamininą. Infect Immun (2005) 73(11):7485–94. doi: 10.1128/iai.73.11.7485-7494.2005
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
54. Lee KH, Choi HJ, Lee MG, Lee JB. Zjadliwy antygen Treponema pallidum 47 kDa reguluje ekspresję cząsteczek adhezji komórkowej i wiązanie limfocytów T z hodowanymi ludzkimi komórkami śródbłonka mikronaczyniowego skóry. Yonsei Med J. (2000) 41(5):623–33. doi: 10.3349/ymj.2000.41.5.623
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
55. Riley BS, Oppenheimer-Marks N, Hansen EJ, Radolf JD, Norgard MV. Zjadliwy treponema pallidum aktywuje ludzkie komórki śródbłonka naczyniowego. J. Infect Dis (1992) 165(3):484–93. doi: 10.1093/infdis/165.3.484
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
56. Zhang RL, Zhang JP, Wang QQ. Rekombinowane białko Treponema pallidum Tp0965 aktywuje komórki śródbłonka i zwiększa przepuszczalność monowarstwy komórek śródbłonka. PloS One (2014) 9(12):e115134. doi: 10.1371/journal.pone.0115134
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
57. Houston S, Russell S, Hof R, Roberts AK, Cullen P, Irvine K i in. Wielofunkcyjna rola związanego z pallilizyną białka T reponema pallidum, Tp0750, w promowaniu fibrynolizy i degradacji składników macierzy zewnątrzkomórkowej. Mol Microbiol (2013) 91(3):618–34. doi: 10.1111/mmi.12482
58. Houston S, Hof R, Francescutti T, Hawkes A, Boulanger MJ, Cameron CE. Dwufunkcyjna rola macierzy zewnątrzkomórkowej treponema pallidum wiążącej adhezynę Tp0751. Infect Immun (2011) 79(3):1386–98. doi: 10.1128/iai.01083-10
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
59. Sprzedaj S, Salman J, Norris SJ. Ponowne zakażenie królików odpornych na wrzód krętkowy krętkiem bladym i. badania świetlne i immunofluorescencyjne. Am J. Pathol (1985) 118(2):248–55.
60. Borenstein LA, Ganz T, Sell S, Lehrer RI, Miller’ AJN. Udział defensyn leukocytów królika w odpowiedzi gospodarza na kiłę eksperymentalną. Infect Immun (1991) 59(4):1368–77. doi: 10.1128/iai.59.4.1368-1377.1991
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
61. Musher DM, Hague-Park M, Gyorkey F, Anderson DC, Baughn RE. Interakcja między treponema pallidum a ludzkimi leukocytami wielojądrzastymi. J. Infect Dis (1983) 147(1):77–86. doi: 10.1093/infdis/147.1.77
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
62. Shin JL, Chung KY, Kang JM, Lee TH, Lee MG. Wpływ Treponema pallidum na ludzkie komórki dendrytyczne. Yonsei Med J. (2004) 45(3):515–22. doi: 10.3349/ymj.2004.45.3.515
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
63. Bouis DA, Popova TG, Takashima A, Norgard MV. Komórki dendrytyczne fagocytują i są aktywowane przez Treponema pallidum. Infect Immun (2001) 69(1):518–28. doi: 10.1128/iai.69.1.518-528.2001
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
64. Sell S, Baker-Zander S, Powell HC. Eksperymentalne kiłowe zapalenie jąder u królików: ultrastrukturalny wygląd treponema pallidum podczas fagocytozy i rozpuszczania przez makrofagi in vivo . Lab Invest (1982) 46(4):355–64.
65. Lukehart SA, Baker-Zander SA, Lloyd RMC, Sell S. Charakterystyka reakcji limfocytów we wczesnej kile eksperymentalnej. II. charakter nacieku komórkowego i rozmieszczenie krętka bladego w zmianach jąder. J. Immunol (1980) 124(1):461–7.
66. Sell S, Baker-Zander SA, Lloyd RM. Rozrost komórek T tkanek limfatycznych królików zakażonych krętkiem bladym: dowód na silną odpowiedź immunologiczną. Sex Transm Dis (1980) 7 (2): 74–84. doi: 10.1097/00007435-198004000-00009
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
67. Lider BT, Godornes C, VanVoorhis WC, Lukehart SA. Limfocyty CD4+ i interferon gamma dominują w miejscowych odpowiedziach immunologicznych we wczesnej kile eksperymentalnej. Infect Immun (2007) 75(6):3021–6. doi: 10.1128/iai.01973-06
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
68. Carlson JA, Dabiri G, Cribier B, Sell S, Physician R. Immunopatobiologia kiły: objawy i przebieg kiły zależą od poziomu nadwrażliwości typu opóźnionego. Am J. Dermatopatol (2011) 33(5):433–60. doi: 10.1097/dad.0b013e3181e8b587
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
69. Sykes JA, Kalan J. Wewnątrzkomórkowy treponema pallidum w komórkach zmiany syfilitycznej szyjki macicy. Am J. Obstet Gynecol (1975) 122(3):361–7. doi: 10.1016/0002-9378(75)90185-4
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
70. Sykes JA, Miller JN. Wewnątrzkomórkowa lokalizacja treponema pallidum (szczep Nichols) w jądrach królika. Infect Immun (1971) 4(3):307–14. doi: 10.1128/iai.4.3.307-314.1971
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
71. Sykes JA, Miller JN, Kalan AJ. Treponema pallidum w komórkach pierwotnego wrzodu u kobiety. Br J. Venereal Dis (1974) 50(1):40–4. doi: 10.1136/sti.50.1.40
72. Van Voorhis WC, Barrett LK, Nasio JM, Plummer FA, Lukehart SA. Zmiany kiły pierwotnej i wtórnej zawierają aktywowane cytolityczne limfocyty T. Infect Immun (1996) 64(3):1048–50. doi: 10.1128/iai.64.3.1048-1050.1996
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
73. Moore MW, Cruz AR, LaVake CJ, Marzo AL, Eggers CH, Salazar JC i in. Fagocytoza Borrelia burgdorferi i Treponema pallidum nasila wrodzoną aktywację immunologiczną i indukuje wytwarzanie interferonu gamma. Infect Immun (2007) 75(4):2046–62. doi: 10.1128/iai.01666-06
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
74. Lukehart SA, Miller JN. Wykazanie fagocytozy in vitro treponema pallidum przez makrofagi otrzewnowe królika. J. Immunol (1978) 121(5):2014–24. doi: 10.4049/jimmunol.121.5.2014
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
75. Baker-Zander SA, Lukehart SA. Zabijanie za pośrednictwem makrofagów opsonizowanego krętka bladego. J. Infect Dis (1992) 165(1):69–74. doi: 10.1093/infdis/165.1.69
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
76. Baker-Zander SA, Shaffer JM, Lukehart SA. Charakterystyka zapotrzebowania na surowicę w celu zabijania za pośrednictwem makrofagów Treponema pallidum ssp. pallidum: Związek z rozwojem przeciwciał opsonizujących. FEMS Immunol Med Microbiol (1993) 6(4):273–9. doi: 10.1111/j.1574-695x.1993.tb00339.x
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
77. Hawley KL, Cruz AR, Benjamin SJ, La Vake CJ, Cervantes JL, LeDoyt M i in. IFNγ wzmaga wzmocnioną CD64 fagocytozę krętka bladego opsonizowanego ludzką surowicą syfilityczną przez ludzkie makrofagi. Immunol (2017) 8:1227. doi: 10.3389/fimmu.2017.01227
78. Chen H., Tong ML, Liu LL, Lin LR, Yang TC. Cały proces interakcji makrofag-krętek blady: fagocytoza opsoniczna, fagocytoza nieopsoniczna i aktywna inwazja. Int Immunopharmacol (2022) 107:108657. doi: 10.1016/j.intimp.2022.108657
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
79. Lin LR, Liu W, Zhu XZ, Chen YY, Gao ZX, Gao K i in. Treponema pallidum promuje polaryzację makrofagów i aktywuje szlak inflammasomu NLRP3 w celu indukcji wytwarzania interleukiny-1β. BMC Immunol (2018) 19(1):28. doi: 10.1186/s12865-018-0265-9
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
80. Lukehart SA, Baker-Zander SA, Sell S. Charakterystyka humoralnej odpowiedzi immunologicznej królika na antygeny krętka bladego po doświadczalnym zakażeniu i terapii. Sex Transm Dis (1986) 13(1): 9–15. doi: 10.1097/00007435-198601000-00003
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
81. Lider BT, Hevner K, Molini BJ, Barrett LK, Van Voorhis WC, Lukehart SA. Odpowiedzi przeciwciał wywołane przeciwko powtórzeniom białek Treponema pallidum różnią się podczas zakażenia różnymi izolatami Treponema pallidum subsp. blady. Infect Immun (2003) 71(10):6054–7. doi: 10.1128/iai.71.10.6054-6057.2003
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
82. Moskophidis M. Analiza humoralnej odpowiedzi immunologicznej na krętek blady w różnych stadiach nieleczonej kiły ludzkiej. Zentralblatt fur Bakteriologie (1989) 271(2):171–9. doi: 10.1016/S0934-8840(89)80070-2
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
83. Fitzgerald TJ. Aktywacja klasycznej i alternatywnej drogi dopełniacza przez treponema pallidum subsp. pallidum i treponema vincentii. Zakażenie i odporność (1987). 55(9):2066–73. doi: 10.1128/iai.55.9.2066-2073.1987
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
84. Blanco DR, Walker EM, Haake DA, Champion CI, Miller JN, Lovett MA. Aktywacja dopełniacza ogranicza szybkość działania krętkobójczego in vitro i koreluje z agregacją za pośrednictwem przeciwciał rzadkiego białka błony zewnętrznej Treponema pallidum. J. Immunol (1990) 144(5):1914–21. doi: 10.4049/jimmunol.144.5.1914
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
85. Ryż M., Fitzgerald TJ. Immobilizacja immunologiczna krętka bladego: ponowne spojrzenie na interakcje przeciwciał i dopełniacza. Can J. Microbiol (1985) 31(12):1147–51. doi: 10,1139/m85-216
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
86. Fearon DT. Regulacja przez błonowy kwas sialowy zależnego od β1H rozpadu-dysocjacji amplifikacyjnej konwertazy C3 alternatywnego szlaku dopełniacza. Proc Natl Acad Sci USA (1978) 75(4):1971–5. doi: 10.1073/pnas.75.4.1971
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
87. Meri S., Pangburnt MK. Rozróżnienie pomiędzy aktywatorami i nieaktywatorami alternatywnej ścieżki dopełniacza: Regulacja poprzez miejsce wiązania kwasu sialowego/polianion na czynniku h. Proc Natl Acad Sci USA (1990) 87(10):3982–6. doi: 10.1073/pnas.87.10.3982
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
88. Ram S, Sharma AK, Simpson SD, Gulati S, Mcquillen DP, Pangburn MK i in. Nowe miejsce wiązania kwasu sialowego na czynniku h pośredniczy w oporności sialilowanej Neisseria gonorrhoeae na surowicę. J Exp Med (1998) 187(5):743–52:Ryż. doi: 10.1084/jem.187.5.743
89. Jarvis GA, Vedros NA. Kwas sialowy z grupy b Neisseria meningitidis reguluje aktywację alternatywnego szlaku dopełniacza. Infect Immun (1987) 55(1):174–80. doi: 10.1128/iai.55.1.174-180.1987
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
90. Sprzedam S, Hsu PL. Opóźniona nadwrażliwość, odchylenia immunologiczne, przetwarzanie antygenów i selekcja podzbioru komórek T w patogenezie kiły i projektowaniu szczepionek. Immunol dzisiaj (1993) 14(12):576–82. doi: 10.1016/0167-5699(93)90196-r
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
91. Biskup NH, Miller JN. Odporność humoralna w kile doświadczalnej. II. związek czynników neutralizujących w surowicy odpornościowej z opornością nabytą. J. Immunol (1976) 117(1):197–207.
92. Biskup NH, Miller JN. Odporność humoralna w kile eksperymentalnej. J. Immunol (1976) 117(1):191–6.
93. Hanff PA, Biskup NH, Miller JN, Lovett MA. Humoralna odpowiedź immunologiczna w kile doświadczalnej na polipeptydy krętka bladego. J. Immunol (1983) 131(4):1973–7. doi: 10.4049/jimmunol.131.4.1973
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
94. Arroll TW, Centurion-Lara A, Lukehart SA, Van Voorhis WC. Odpowiedzi komórek T na treponema pallidum subsp. pallidum w przebiegu doświadczalnego zakażenia kiłą. Infect Immun (1999) 67(9):4757–63. doi: 10.1128/IAI.67.9.4757-4763.1999
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
95. Sell S, Gamboa D, Baker-Zander SA, Lukehart SA, Miller JN. Odpowiedź gospodarza na treponema pallidum u królików zakażonych śródskórnie: dowody na utrzymywanie się infekcji w miejscach lokalnych i odległych. J Invest Dermatol (1980) 75(6):470–5. doi: 10.1111/1523-1747.ep12524230
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
96. Yogeswari L., Chacko CW. Trwałość t. pallidum i jego znaczenie w seropozytywnej kile późnej leczonej penicyliną. Br. J. Vener Dis (1971) 47(5):339–47. doi: 10.1136/sti.47.5.339
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
97. Wicher K, Abbruscato F, Wicher V, Collins DN, Auger I, Horowitz HW. Identyfikacja uporczywej infekcji w kile eksperymentalnej metodą PCR. Infect Immun (1998) 66(6):2509–13. doi: 10.1128/IAI.66.6.2509-2513.1998
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
98. Castro R, Prieto E, Águas MJ, Manata MJ, Botas J, Santo I i in. Wykrywanie DNA Treponema pallidum sp pallidum w kile utajonej. Int J STD AIDS (2007) 18(12):842–5. doi: 10.1258/095646207782716901
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
99. Engelkens HJ, Ten Kate FJ, Judanarso J, Vuzevski VD, Van Lier JB, Godschalk JC i in. Lokalizacja krętków i charakterystyka nacieku zapalnego w biopsjach skóry pobranych od pacjentów z kiłą pierwotną lub wtórną lub wczesną chorobą zakaźną. Genitourin Med (1993) 69(2):102–7. doi: 10.1136/sti.69.2.102
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
100. Singh R, Teranno D. Niezwykły objaw wysypki skórnej. J. Clin Pathol (2017) 70(12):1088. doi: 10.1136/jclinpath-2016-204007
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
101. Alessi E, Innocenti M, Ragusa G. Kiła wtórna. morfologia kliniczna i histopatologia. Am J. Dermatopatol (1983) 5(1):11–7. doi: 10.1097/00000372-198302000-00004
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
102. Baker-Zander SA, Roddy RE, Handsfield HH, Lukehart SA. Reaktywność przeciwciał IgG i IgM na antygeny krętka bladego po leczeniu kiły. Sex Transm Dis (1986) 13(4):214–20. doi: 10.1097/00007435-198610000-00002
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
103. Bryza AC. Choroby zakaźne płodu i noworodka, wyd. 6. Arch Dis Dziecko Płodowe Noworodkowe Ed (2007) 92:F156. doi: 10.1136/adc.2006.102566
104. Baughn RE, Jorizzo JL, Adams CB, Musher DM. Reakcja klasy Ig i podklasy IgG na krętek blady u pacjentów z kiłą. J Clin Immunol (1988) 8(2):128–39. doi: 10.1007/bf00917901
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
105. Liu LL, Chao PL, Zhang HL, Tong ML, Liu GL, Lin LR i in. Analiza podzbiorów limfocytów u pacjentów z kiłą układu nerwowego HIV-ujemnego. Diagnoza Microbiol Infect Dis (2013) 75(2):165–8. doi: 10.1016/j.diagmicrobio.2012.10.007
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
106. Podwinska J, Lusiak M, Zaba R, Bowszyc J. Wzór i poziom cytokin wydzielanych przez limfocyty Th1 i Th2 u pacjentów z kiłą korelują z postępem choroby. FEMS Immunol Med Microbiol (2000) 28(1):1–14. doi: 10.1111/j.1574-695x.2000.tb01451.x
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
107.Miller JN. Odporność na kiłę eksperymentalną. VI. skuteczne szczepienie królików treponema pallidum, szczepem Nicholsa, osłabionym przez napromienianie. J. Immunol (1973) 110(5):1206–15. doi: 10.4049/jimmunol.110.6.np
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
108. Magnuson HJ, Thomas EW, Olansky S, Kaplan BI, de Mello L, Cutler JC. Kiła inokulacyjna u ochotników. Medycyna (1956) 35(1):33–82. doi: 10.1097/00005792-195602000-00002
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
109. Marra CM, Maxwell CL, Sahi SK, Tantalo LC, Dunaway SB, Lukehart SA. Kiła przebyta zmienia przebieg kolejnych epizodów kiły. Clin Infect Dis (2022) 74: e1–5. doi: 10.1093/cid/ciab287
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
110. Cullen PA, Haake DA, Adler B. Białka błony zewnętrznej patogennych krętków. FEMS Microbiol Rev (2004) 28:291–318. doi: 10.1016/j.femsre.2003.10.004
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
111. Hawley KL, Montezuma-Rusca JM, Delgado KN, Singh N, Uversky VN, Caimano MJ i in. Modelowanie strukturalne repertuaru białek błony zewnętrznej treponema pallidum: mapa drogowa dekonwolucji patogenezy kiły i opracowania szczepionki przeciw kiły. J Bacteriol (2021) 203: 82–103. doi: 10.1128/JB
112. Molini B, Fernandez MC, Godornes C, Vorobieva A, Lukehart SA, Giacani L. Mapowanie epitopów komórek B wariantów TprC i TprD podgatunku treponema pallidum stanowi podstawę do opracowania szczepionki przeciwko ludzkim treponematozom. Przód Immunol (2022) 13:862491/bibtex. doi: 10.3389/fimmu.2022.862491/bibtex
113. Kojima N, Konda KA, Klausner JD. Uwagi na temat rozwoju szczepionki przeciwko kiły. Przedni Immunol (2022) 13:952284. doi: 10.3389/fimmu.2022.952284
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
114. Romeis E, Tantalo L, Lieberman N, Phung Q, Greninger A, Giacani L. Inżynieria genetyczna treponema pallidum subsp. pallidum, krętek kiły. PloS Pathog (2021) 17(7):e1009612. doi: 10.1371/journal.ppat.1009612
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
115. Centurion-Lara A, Castro C, Barrett L, Cameron C, Mostowfi M, Van Voorhis WC i in. Homolog głównego białka osłonki Treponema pallidum tpr K jest celem przeciwciał opsonicznych i ochronnej odpowiedzi immunologicznej. J Exp Med (1999) 189(4):647–56. doi: 10.1084/jem.189.4.647
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
116. Edwards AM, Jenkinson HF, Woodward MJ, Dymock D. Właściwości wiążące i regiony pośredniczące w adhezji głównego białka osłonki krętka denticola ATCC 35405. Infect Immun (2005) 73(5):2891–8. doi: 10.1128/iai.73.5.2891-2898.2005
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
117. Fenno JC, Müller KH, McBride BC. Analiza sekwencji, ekspresja i aktywność wiązania rekombinowanego głównego białka osłonki zewnętrznej (Msp) krętka denticola. J. Bacteriol (1996) 178(9):2489–97. doi: 10.1128/jb.178.9.2489-2497.1996
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
118. Mathers DA, Leung WK, Fenno JC, Hong Y, McBride BC. Główny kompleks białek powierzchniowych treponema denticola depolaryzuje i indukuje kanały jonowe w błonach komórkowych HeLa. Infect Immun (1996) 64(8):2904–10. doi: 10.1128/iai.64.8.2904-2910.1996
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
119. Anand A, Luthra A, Dunham-Ems S, Caimano MJ, Karanian C, LeDoyt M i in. TprC/D (Tp0117/131), trimeryczne, tworzące pory rzadkie białko błony zewnętrznej krętka pallidum, ma dwudzielną strukturę domeny. J Bacteriol (2012) 194(2):2321–33. doi: 10.1128/jb.00101-12
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
120. Anand A, Ledoyt M, Karanian C, Luthra A, Koszelak-Rosenblum M, Malkowski MG i in. Dwuczęściowa topologia powtórzeń białek Treponema pallidum C/D i I: Insercja błony zewnętrznej, trimeryzacja i funkcja poriny wymagają domeny c-końcowej-Barrel. J Biol Chem (2015) 290(19):12313–31. doi: 10.1074/jbc.m114.629188
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
121. Giacani L, Sambri V, Marangoni A, Cavrini F, Storni E, Donati M i in. Ocena immunologiczna i analiza lokalizacji komórkowej antygenu TprI treponema pallidum subsp. blady. Infect Immun (2005) 73(6):3817–22. doi: 10.1128/iai.73.6.3817-3822.2005
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
122. Giacani L, Molini B, Godornes C, Barrett L, Van Voorhis W, Centurion-Lara A i in. Ilościowa analiza ekspresji genu tpr w izolatach treponema pallidum: różnice między izolatami i korelacja z reakcją limfocytów T w kile eksperymentalnej. Infect Immun (2007) 75(1):104–12. doi: 10.1128/iai.01124-06
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
123. Giacani L, Godornes C, Puray-Chavez M, Guerra-Giraldez C, Tompa M, Lukehart SA i in. TP0262 jest modulatorem aktywności promotora genów podrodziny II tpr treponema pallidum ssp. blady. Mol Microbiol (2009) 72(5):1087–99. doi: 10.1111/j.1365-2958.2009.06712.x
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
124. Giacani L, Lukehart S, Centurion-Lara A. Długość homopolimerowych powtórzeń guanozyny moduluje aktywność promotora genów tpr podrodziny II treponema pallidum ssp. blady. FEMS Immunol Med Microbiol (2007) 51(2):289–301. doi: 10.1111/j.1574-695X.2007.00303.x
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
125. Haynes AM, Fernandez M, Romeis E, Mitjà O, Konda KA, Vargas SK i in. Analiza transkrypcyjna i immunologiczna domniemanego białka błony zewnętrznej i kandydata na szczepionkę TprL krętka bladego. PloS Negl Trop Dis (2021) 15(1):e0008812. doi: 10.1371/journal.pntd.0008812
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
126. Lian T, Zhang B, Giacani L, Kou C, Yang X, Zhang R i in. Pełnej długości TprK treponema pallidum subsp. pallidum w nanodyskach lipidowych to monomeryczna porina. Enzym Microb Technol (2022) 153:109897. doi: 10.1016/j.enzmictec.2021.109897
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
127. Centurion-Lara A, Godornes C, Castro C, Van Voorhis WC, Lukehart SA. Gen tprK jest heterogenny wśród szczepów Treponema pallidum i ma wiele alleli. Infect Immun (2000) 68(2):824–31. doi: 10.1128/IAI.68.2.824-831.2000
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
128. Addetia A, Lin MJ, Phung Q, Xie H, Huang ML, Ciccarese G i in. Ocena różnorodności tprk pełnej długości u treponema pallidum subsp. blady. mBio (2020) 11(5):1–19. doi: 10.1128/mbio.02726-20
129. Lafond RE, Centurion-lara A, Godornes C, Van Voorhis WC, Lukehart SA. Różnorodność sekwencji TprK kumuluje się podczas infekcji królików Treponema pallidum subsp. szczep pallidum nichols. Infekcja i odporność (2006) 74(3):1896–906. doi: 10.1128/iai.74.3.1896
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
130. Giacani L, Molini BJ, Kim EY, Godornes BC, Leader BT, Tantalo LC i in. Zmienność antygenowa treponema pallidum: różnorodność sekwencji TprK gromadzi się w odpowiedzi na presję immunologiczną podczas kiły eksperymentalnej. J. Immunol (2010) 184(7):3822–9. doi: 10.4049/jimmunol.0902788
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
131. Lin MJ, Haynes AM, Addetia A, Lieberman NAP, Phung Q, Xie H i in. Podłużne profilowanie TprK treponema pallidum subsp. rozmnażanego in vivo i in vitro . pallidum ujawnia akumulację wariantów antygenowych przy braku ciśnienia immunologicznego. PloS Negl Trop Dis (2021) 15(9):e0009753. doi: 10.1371/JOURNAL.PNTD.0009753
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
132. Morgan CA, Lukehart SA, Van Voorhis WC. Ochrona przed kiłą koreluje ze specyficznością przeciwciał przeciwko regionom zmiennym białka powtórzeń krętka pallidum K. Infect Immun (2003) 71(10):5605–12. doi: 10.1128/iai.71.10.5605-5612.2003
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
133. Morgan CA, Molini BJ, Lukehart SA, Van Voorhis WC. Segregacja epitopów komórek b i T białka K powtórzeń krętka pallidum do regionów zmiennych i konserwatywnych podczas eksperymentalnej infekcji kiłą. J. Immunol (2002) 169(2):952–7. doi: 10.4049/jimmunol.169.2.952
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
134. Morgan CA, Lukehart SA, Van Voorhis WC. Immunizacja n-końcową częścią białka K powtórzeń krętka pallidum osłabia rozwój zmian syfilitycznych w modelu królika. Infect Immun (2002) 70(12):6811. doi: 10.1128/IAI.70.12.6811-6816.2002
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
135. Hazlett KR, Sellati TJ, Nguyen TT, Cox DL, Clawson ML, Caimano MJ i in. Białko TprK treponema pallidum jest periplazmatyczne i nie jest celem przeciwciał opsonicznych ani odporności ochronnej. J Exp Med (2001) 193(9):1015–26. doi: 10.1084/jem.193.9.1015
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
136. Sun ES, Molini BJ, Barrett LK, Centurion-Lara A, Lukehart SA, Van Voorhis WC. Podrodzina I Rodzina białek powtórzeń Treponema pallidum: zmienność sekwencji i odporność. Zarażają drobnoustroje (2004) 6: 725–37. doi: 10.1016/j.micinf.2004.04.001
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
137. Nandi B, Nandy RK, Sarkar A, Ghose AC. Cechy strukturalne, właściwości i regulacja białka błony zewnętrznej W (OmpW) vibrio cholerae. Mikrobiologia (2005) 151:2975–86. doi: 10.1099/mik.0.27995-0
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
138. Hu WS, Li PC, Cheng CY. Korelacja między opornością na ceftriakson salmonelli enterica serovar typhimurium a ekspresją białek błony zewnętrznej OmpW i białka podobnego do Ail/OmpX, które są regulowane przez BaeR układu dwuskładnikowego. Środki przeciwdrobnoustrojowe Chemother (2005) 49(9):3955–8. doi: 10.1128/aac.49.9.3955-3958.2005
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
139. Hong H, Patel DR, Tamm LK, Van Den Berg B. Białko błony zewnętrznej OmpW tworzy ośmioniciową beczkę beta z kanałem hydrofobowym. J Biol Chem (2006) 281(11):7568–77. doi: 10.1074/jbc.m512365200
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
140. Giacani L, Brandt SL, Ke W, Reid TB, Molini BJ, Iverson-Cabral S i in. Transkrypcja TP0126, treponema pallidum domniemanego homologu OmpW, jest regulowana przez długość homopolimerowego powtórzenia guanozyny. Infect Immun (2015) 83(6):2275–89. doi: 10.1128/iai.00360-15
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
141. Haynes AM, Godornes C, Ke W, Giacani L. Ocena zdolności ochronnej treponema pallidum subsp. pallidum Tp0126 Homolog OmpW w króliczym modelu kiły. Infect Immun (2019) 87(8):e00323–19. doi: 10.1128/IAI.00323-19
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
142. Cox DL, Radolf JD. Wstawienie fluorescencyjnych sond kwasów tłuszczowych do zewnętrznych błon chorobotwórczych krętków treponema pallidum i Borrelia burgdorferi. Mikrobiologia (2001) 147: 1161–9. doi: 10.1099/00221287-147-5-1161
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
143. Delgado KN, Montezuma-Rusca JM, Orbe IC, Caimano MJ, La Vake CJ, Luthra A i in. Pętle zewnątrzkomórkowe treponema pallidum ortologów FadL TP0856 i TP0858 wywołują przeciwciała IgG i komórki b specyficzne dla IgG+ w króliczym modelu kiły eksperymentalnej. mBio (2022) 13(4):e0163922. doi: 10.1128/mbio.01639-22
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
144. Bleuel C, Große C, Taudte N, Scherer J, Wesenberg D, Krauß GJ i in. TolC bierze udział w wypływie enterobaktyny przez zewnętrzną błonę escherichia coli. J Bacteriol (2005) 187(19):6701–7. doi: 10.1128/jb.187.19.6701-6707.2005
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
145. Brautigam CA, Deka RK, Ouyang Z, Machius M, Knutsen G, Tomchick DR i in. Analizy biofizyczne i bioinformatyczne wskazują na udział lipoproteiny treponema pallidum tp34 (tp0971) w homeostazie metali przejściowych. J Bacteriol (2012) 194(24):6771–81. doi: 10.1128/JB.01494-12
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
146. Kubanov A, Runina A, Deryabin D. Nowe rekombinowane antygeny treponema pallidum do diagnostyki kiły: stan obecny i perspektywy na przyszłość. BioMed Res Int (2017) 2017:1436080. doi: 10.1155/2017/1436080
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
147. Blanco DR, Champion CI, Exner MM, Shang ES, Skare JT, Hancock REW i in. Rekombinowane rzadkie białko błony zewnętrznej 1 (Tromp1) treponema pallidum ulegające ekspresji w escherichia coli ma aktywność porową i ekspozycję na antygeny powierzchniowe. J. Bacteriol (1996) 178(23):6685–92. doi: 10.1128/jb.178.23.6685-6692.1996
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
148. Deka RK, Lee YH, Hagman KE, Shevchenko D, Lingwood CA, Hasemann CA i in. Fizykochemiczne dowody na to, że Treponema pallidum TroA jest metaloproteiną zawierającą cynk i pozbawioną struktury przypominającej porinę. J. Bacteriol (1999) 181(14):4420–3. doi: 10.1128/jb.181.14.4420-4423.1999
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
149. Akins DR, Robinson E, Szewczenko D, Elkins C, Cox DL, Radolf JD. Tromp1, domniemane rzadkie białko błony zewnętrznej, jest zakotwiczone przez nierozszczepioną sekwencję sygnałową do błony cytoplazmatycznej krętka bladego. J. Bacteriol (1997) 179(16):5076–86. doi: 10.1128/jb.179.16.5076-5086.1997
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
150. Hardham JM, Stamm LV, Porcella SF, Frye JG, Barnes NY, Howell JK i in. Identyfikacja i analiza transkrypcji operonu Treponema pallidum kodującego przypuszczalny system transportu ABC, homolog białka represora aktywowanego żelazem i homolog enzymu szlaku glikolitycznego. Gene (1997) 197(1):47–64. doi: 10.1016/S0378-1119(97)00234-5
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
151. Xu M, Xie Y, Jiang C, Xiao Y, Kuang X, Zhao F i in. Nowatorski test ELISA wykorzystujący rekombinowane białko błony zewnętrznej rTp0663 jako antygen do diagnostyki serologicznej kiły. Int J Infect Dis (2016) 43:51–7. doi: 10.1016/j.ijid.2015.12.013
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
152. Xu M, Xie Y, Zheng K, Luo H, Tan M, Zhao F i in. Dwóch potencjalnych kandydatów na szczepionkę przeciwko kile hamuje rozprzestrzenianie się krętka bladego. Przedni Immunol (2021) 12:759474. doi: 10.3389/fimmu.2021.759474
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
153. Djokic V, Giacani L, Parveen N. Analiza specyficzności wiązania komórek gospodarza za pośrednictwem adhezyny Tp0136 czynnika kiły treponema pallidum subsp. blady. PloS Negl Trop Dis (2019) 13(5):e0007401. doi: 10.1371/journal.pntd.0007401
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
154. Xu QY, Wang YJ, Lin LR, Liu LL, Yang TC. Lipoproteina błony zewnętrznej Tp0136 stymuluje aktywację i agregację ludzkich płytek krwi poprzez PAR1 w celu wzmocnienia sygnalizacji Gq/Gi. Przedni Immunol (2022) 13:818151. doi: 10.3389/fimmu.2022.818151
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
155. Cameron CE, Brown EL, Kuroiwa JMY, Schnapp LM, Brouwer NL. Białka wiążące fibronektynę Treponema pallidum. Journal of Bacteriology (2004) 186(20):7019–22. doi: 10.1128/jb.186.20.7019
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
156. Bamford CV, Francescutti T, Cameron CE, Jenkinson HF, Dymock D. Charakterystyka nowej rodziny białek wiążących fibronektynę z domenami peptydazy M23 z Treponema denticola. Mol Oral Microbiol (2010) 25(6):369–83. doi: 10.1111/j.2041-1014.2010.00584.x
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
157. Tomson FL, Conley PG, Norgard MV, Hagman KE. Ocena ekspozycji na powierzchnię komórki i potencjału szczepionkowego białek błony zewnętrznej będących kandydatami na krętek blady. Zarażanie drobnoustrojami (2007) 9(11):1267–75. doi: 10.1016/j.micinf.2007.05.018
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
158. Van Voorhis WC, Barrett LK, Lukehart SA, Schmidt B, Schriefer M, Cameron CE. Serodiagnostyka kiły: Przeciwciała przeciwko rekombinowanym białkom Tp0453, Tp92 i gpd są czułymi i swoistymi wskaźnikami zakażenia krętkiem bladym. J Clin Microbiol (2003) 41(8):3668. doi: 10.1128/jcm.41.8.3668-3674.2003
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
159. Chan K, Nasereddin T, Alter L, Centurion-Lara A, Giacani L, Parveen N. Lipoproteina Treponema pallidum TP0435 wyrażana w borrelia burgdorferi wytwarza wiele izoform powierzchniowych/peryplazmatycznych i pośredniczy w przyleganiu. Science Rep (2016) 6: 1–13. doi: 10.1038/srep25593
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
160. Cox DL, Akins DR, Porcella SF, Norgard MV, Radolf JD. Treponema pallidum w mikrokropelkach żelowych: nowatorska strategia badania architektury molekularnej krętkowej. Mol Microbiol (1995) 15(6):1151–64. doi: 10.1111/j.1365-2958.1995.tb02288.x
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
161. Brautigam CA, Deka RK, Liu WZ, Norgard MV. Wgląd w potencjalną funkcję i organizację błony lipoproteiny TP0435 (Tp17) z krętka bladego uzyskany z analiz strukturalnych i biofizycznych. Białko Sci (2015) 24(1):11–9. doi: 10.1002/pro.2576
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
162. Parveen N, Fernandez MC, Haynes AM, Zhang RL, Godornes BC, Centurion-Lara A i in. Niepatogenna Borrelia burgdorferi eksprymująca antygeny Treponema pallidum TprK i Tp0435 jako nowe podejście do oceny kandydatów na szczepionkę przeciw kiły. Szczepionka (2019). 37(13):1807–18. doi: 10.1016/j.vaccine.2019.02.022
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
163. Cameron CE. Identyfikacja białka wiążącego lamininę Treponema pallidum. Infect Immun (2003) 71(5):2525–33. doi: 10.1128/iai.71.5.2525-2533.2003
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
164. Dickerson MT, Abney MB, Cameron CE, Knecht M, Bachas LG, Anderson KW. Wiązanie fibronektyny z fragmentem białka adhezyny treponema pallidum rTp0483 na funkcjonalizowanych samoorganizujących się monowarstwach. Bioconjug Chem (2012) 23(2):184–95. doi: 10.1021/bc200436x
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
165. Houston S, Taylor JS, Denchev Y, Hof R, Zuerner RL, Cameron CE. Konserwacja białek oddziałujących z gospodarzem Tp0750 i palilizyny wśród treponemów i ograniczenie zdolności proteolitycznej krętka bladego. Infect Immun (2015) 83(11):4204–16. doi: 10.1128/iai.00643-15
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
166. Luthra A, Montezuma-Rusca JM, la Vake CJ, LeDoyt M, Delgado KN, Davenport TC i in. Dowody na to, że immunizacja TP0751, dwuczęściową lipoproteiną Treponema pallidum z wewnętrznie nieuporządkowanym regionem i fałdem lipokalinowym, nie chroni w królikim modelu doświadczalnej kiły. PloS Pathog (2020) 16(9):e1008871. doi: 10.1371/journal.ppat.1008871
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
167. Kao WCA, Pětrošová H, Ebady R, Lithgow KV, Rojas P, Zhang Y i in. Identyfikacja Tp0751 (Pallilysin) jako adhezyny naczyniowej Treponema pallidum poprzez heterologiczną ekspresję w krętku choroby z Lyme. Sci Rep (2017) 7(1):1–13. doi: 10.1038/s41598-017-01589-4
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
168. Lithgow KV, Hof R, Wetherell C, Phillips D, Houston S, Cameron CE. Zdefiniowany kandydat na szczepionkę przeciwko kile hamuje rozprzestrzenianie się podgatunku Treponema pallidum pallidum. Nat Commun (2017) 8(1):1–10. doi: 10.1038/ncomms14273
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
169. Primus S, Rocha SC, Giacani L, Parveen N. Identyfikacja i ocena funkcjonalna pierwszej adhezyny łożyskowej krętka bladego, która może odgrywać kluczową rolę w kile wrodzonej. Przedni Mikrobiol (2020) 11:621654. doi: 10.3389/FMICB.2020.621654
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
170. Giri TK, Tollefsen DM. Łożyskowy siarczan dermatanu: izolacja, działanie przeciwzakrzepowe i związek z kofaktorem heparyny II. Krew (2006) 107(7):2753. doi: 10.1182/blood-2005-09-3755
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
171. Suga N, Sugimura M, Koshiishi T, Yorifuji T, Makino S, Takeda S. Heparyna/siarczan heparanu/CD44-v3 zwiększa migrację komórek w terminowych unieśmiertelnionych ludzkich komórkach trofoblastu pochodzących z łożyska. Biol Reprod (2012) 86(5):134, 1–8. doi: 10.1095/biolreprod.111.093690
172. Lala PK, Nandi P. Mechanizmy migracji trofoblastów, angiogeneza endometrium w stanie przedrzucawkowym: rola dekoryny. Komórka Adh Migr (2016) 10(1-2):111–25. doi: 10.1080/19336918.2015.1106669
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
173. de Oliveira GB, do Vale AM, dos Santos AC, de Moura CEB, Rocha HA de O, de Oliveira MF. Skład i znaczenie glikozaminoglikanów w macicy i łożysku ssaków. Braz Arch Biol Technol (2015) 58(4):512–20. doi: 10.1590/S1516-8913201500281
174. Luthra A, Anand A, Hawley KL, LeDoyt M, La Vake CJ, Caimano MJ i in. Model homologii ujawnia nowe cechy strukturalne i immunodominujący cel powierzchniowy Loop/Opsonic w ortologu Treponema pallidum BamA TP_0326. J Bacteriol (2015) 197(11):1906–20. doi: 10.1128/jb.00086-15
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
175. Desrosiers DC, Anand A, Luthra A, Dunham-Ems SM, Ledoyt M, Cummings MAD i in. TP0326, maszyna do składania β-beczki treponema pallidum a (BamA) ortolog i rzadkie białko błony zewnętrznej. Mol Microbiol (2011) 80(6):1496–515. doi: 10.1111/j.1365-2958.2011.07662.x
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
176. Cameron CE, Lukehart SA, Castro C, Molini B, Godornes C, Van Voorhis WC. Potencjał opsoniczny, zdolność ochronna i zachowanie sekwencji podgatunku treponema pallidum pallidum Tp92. J Infect Dis (2000) 181(4):1401–13. doi: 10.1086/315399
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
177. Stebeck CE, Shaffer JM, Arroll TW, Lukehart SA, Van Voorhis WC. Identyfikacja treponema pallidum subsp. pallidum glicerofosfodiestrowy homolog fosfodiesterazy. FEMS Microbiol Lett (1997) 154(2):303–10. doi: 10.1111/J.1574-6968.1997.TB12660.X
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
178. Cameron CE, Castro C, Lukehart SA, Van Voorhis WC. Funkcja i zdolność ochronna treponema pallidum subsp. pallidum glicerofosfodiestrowa fosfodiesteraza. Infekcja i odporność (1998) 66(12):5763–70. doi: 10.1128/IAI.66.12.5763-5770.1998
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
179. Szewczenko DV, Akins DR, Robinson EJ, Li M, Szewczenko OV, Radolf JD. Identyfikacja homologów tioredoksyny, izomerazy peptydyloprolilo cis-trans i fosfodiesterazy glicerofosfodiestrowej we frakcjach błony zewnętrznej krętka pallidum, krętka kiły. Infect Immun (1997) 65(10):4179–89. doi: 10.1128/iai.65.10.4179-4189.1997
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
180. Szewczenko DV, Sellati TJ, Cox DL, Szewczenko OV, Robinson EJ, Radolf JD. Topologia błony i lokalizacja komórkowa ortologa fosfodiesterazy glicerofosfodiestrowej treponema pallidum (GlpQ). Infect Immun (1999) 67(5):2266. doi: 10.1128/iai.67.5.2266-2276.1999
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
181. Zhao F, Wang S, Zhang X, Gu W, Yu J, Liu S i in. Skuteczność ochronna szczepionki DNA gpd Treponema pallidum wektorowanej nanocząsteczkami chitozanu i połączonej z interleukiną-2. Can J Microbiol (2012) 123: 117–23. doi: 10.1139/w11-115
182. Zhang X, Zhao T, Zeng T, Wu N, Xiao Y, Liu S i in. Domięśniowa pierwotna immunizacja szczepionką zawierającą kwas nukleinowy pcDNA/Gpd-IL-2 i wzmocniona immunizacja adiuwantem błony śluzowej CpG-ODN i rekombinowanym białkiem gpd-IL-2 skutecznie indukowały silną odpowiedź immunologiczną błony śluzowej i działanie immunologiczne chroniące przed zakażeniem skóry krętkiem bladym. Exp Ther Med (2018) 15(3):2533. doi: 10.3892/etm.2018.5689
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
183. Cameron CE, Castro C, Lukehart SA, Van Voorhis WC. Konserwacja sekwencji fosfodiesterazy glicerofosfodiestrowej wśród szczepów Treponema pallidum. Infect Immun (1999) 67(6):3168. doi: 10.1128/iai.67.6.3168-3170.1999
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
184. Thumiger A, Polenghi A, Papinutto E, Battistutta R, Montecucco C, Zanotti G. Struktura krystaliczna antygenu TpF1 z treponema pallidum. Białka: Funkcja struktury Genet (2006) 62(3):827–30. doi: 10.1002/prot.20828
185. Radolf JD, Fehniger TE, Silverblatt FJ, Miller JN, Lovett MA. Powierzchnia zjadliwego treponema pallidum: oporność na wiązanie przeciwciał przy braku dopełniacza i powierzchniowego połączenia rekombinowanego antygenu 4D. Infect Immun (1986) 52(2):579–85. doi: 10.1128/iai.52.2.579-585.1986
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
186. Babolin C, Amedei A, Ozoliņš D, Žileviča A, D’Elios MM, de Bernard M. TpF1 z Treponema pallidum aktywuje inflammasom i promuje rozwój regulatorowych limfocytów T. J. Immunol (2011) 187(3):1377–84. doi: 10.4049/jimmunol.1100615
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
187. Lu DP, Jia J, Wei SF, Zhang WL, Liang R, Liu T i in. Antygen TpF1 Treponema pallidum (kiła) indukuje aktywację makrofagów i przyspiesza indukowane przez P2X7R zależne od NLRP3 uwalnianie IL-1β. Endocr Metab Docelowe leki na zaburzenia immunologiczne (2022) 22(4):425–32. doi: 10.2174/1871530321666211015091109
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
188. McGill MA, Edmondson DG, Carroll JA, Cook RG, Orkiszewski RS, Norris SJ. Charakterystyka i analiza serologiczna proteomu treponema pallidum. Infect Immun (2010) 78(6):2631–43. doi: 10.1128/iai.00173-10
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
189. Pozzobon T, Facchinello N, Bossi F, Capitani N, Benagiano M, Di Benedetto G i in. Antygen Treponema pallidum (kiła) TpF1 indukuje angiogenezę poprzez aktywację szlaku IL-8. Science Rep (2016) 6: 1–14. doi: 10.1038/srep18785
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
190. Cohen SE, Chew Ng RA, Katz KA, Bernstein KT, Samuel MC, Kerndt PR i in. Kiła powtarzająca się wśród mężczyzn uprawiających seks z mężczyznami w Kalifornii w latach 2002-2006: implikacje dla wysiłków na rzecz eliminacji kiły. Am J. Zdrowie publiczne (2012) 102(1):2002–6. doi: 10.2105/ajph.2011.300383
191. de Almeida VC, Donalisio MR, Cordeiro R. Czynniki związane z ponownym zakażeniem kiłą w ośrodkach referencyjnych ds. zakażeń przenoszonych drogą płciową. Rev Saude Publica (2017) 51:64. doi: 10.1590/S1518-8787.2017051006432
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
192. Jain J, Santos GM, Scheer S, Gibson S, Crouch PC, Kohn R i in. Częstość i korelaty ponownego zakażenia kiłą u mężczyzn uprawiających seks z mężczyznami. Zdrowie LGBT (2017) 4(3):232–6. doi: 10.1089/lgbt.2016.0095
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
193. Van der Ende A, Hopman CTP, Zaat S, Essink BBO, Berkhout B, Dankert J. Zmienna ekspresja białka błony zewnętrznej klasy 1 u Neisseria meningitidis jest spowodowana różnicą w odstępach między regionami -10 i -35 promotor. J. Bacteriol (1995) 177(9):2475–80. doi: 10.1128/jb.177.9.2475-2480.1995
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
194. Saunders NJ, Peden JF, Hood DW, Moxon ER. Prosta sekwencja powtarza się w genomie Helicobacter pylori. Mol Microbiol (1998) 27(6):1091–8. doi: 10.1046/j.1365-2958.1998.00768.x
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
195. Van Der Woude MW, Bäumler AJ. Zmienność fazowa i antygenowa bakterii. Clin Microbiol Rev (2004) 17(3):581–611. doi: 10.1128/cmr.17.3.581-611.2004
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
196. Osbak KK, Houston S, Lithgow KV, Meehan CJ, Strouhal M, Šmajs D i in. Charakterystyka wywołującego kiłę Treponema pallidum ssp. pallidum przy użyciu komplementarnej spektrometrii mas. PloS Negl Trop Dis (2016) 10(9):e0004988. doi: 10.1371/journal.pntd.0004988
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
197. Bonifield HR, Hughes KT. Zmienność fazy wici u Salmonella enterica odbywa się za pośrednictwem mechanizmu kontroli potranskrypcyjnej. J Bacteriol (2003) 185(12):3567–74. doi: 10.1128/jb.185.12.3567-3574.2003
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
198. Mouquet H, Scheid JF, Zoller MJ, Krogsgaard M, Ott RG, Shukair S i in. Polireaktywność zwiększa pozorne powinowactwo przeciwciał anty-HIV poprzez heteroligację. Natura (2010) 467(7315):591–5. doi: 10.1038/natura09385
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
199. Wardemann H, Yurasov S, Schaefer A, Young JW, Meffre E, Nussenzweig MC. Dominująca produkcja autoprzeciwciał przez wczesne ludzkie prekursory komórek b. Nauka (2003) 301(5638):1374–7. doi: 10.1126/science.1086907
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
200. Birge RB, Boeltz S, Kumar S, Carlson J, Wanderley J, Calianese D i in. Fosfatydyloseryna jest globalnym sygnałem immunosupresyjnym w eferocytozie, chorobach zakaźnych i nowotworach. Różnicowanie śmierci komórek (2016) 23(6):962–78. doi: 10.1038/cdd.2016.11
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
201. Treede I, Braun A, Sparla R, Kühnel M, Giese T, Turner JR i in. Przeciwzapalne działanie fosfatydylocholiny. J Biol Chem (2007) 282(37):27155–64. doi: 10.1074/jbc.m704408200
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
202. Macek B, Forchhammer K, Hardouin J, Weber-Ban E, Grangeasse C, Mijakovic I. Białkowe modyfikacje potranslacyjne w bakteriach. Nat Rev Microbiol (2019) 17(11):651–64. doi: 10.1038/s41579-019-0243-0
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
203. Sobocińska J, Roszczenko-Jasińska P, Ciesielska A, Kwiatkowska K. Palmitoilacja białek i jej rola w infekcjach bakteryjnych i wirusowych. Przedni Immunol (2018) 8:2003. doi: 10.3389/fimmu.2017.02003
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
204. Kumar S, Caimano MJ, Anand A, Dey A, Hawley KL, LeDoyt ME i in. Zmienność sekwencji rzadkich domen β-baryłki białka błony zewnętrznej w szczepach klinicznych zapewnia wgląd w ewolucję treponema pallidum subsp. pallidum, krętek kiły. mBio (2018) 9(3):e01006–18. doi: 10.1128/mBio.01006-18
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
205. Centurion-Lara A, Giacani L, Godornes C, Molini BJ, Brinck Reid T, Lukehart SA. Dokładna analiza różnorodności genetycznej rodziny genów tpr wśród gatunków, podgatunków i szczepów krętków. PloS Negl Trop Dis (2013) 7(5):e2222. doi: 10.1371/journal.pntd.0002222
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
206. Hawley K, Luthra A, La VC, Ledoyt M, Chen W, Heping Z i in. O01.2 Genetyczna, strukturalna i powierzchniowa zmienność antygenowa OMPeome treponema pallidum: kroki w kierunku globalnej szczepionki przeciw kiły. Infekcja transseksualna (2019) 95:A37–7. doi: 10.1136/sextrans-2019-sti.105
207. Grillová L, Oppelt J, Mikalová L, Nováková M, Giacani L, Niesnerová A i in. Bezpośrednio sekwencjonowane genomy współczesnych szczepów kiły ujawniają napędzaną rekombinacją różnorodność genów kodujących przewidywane antygeny eksponowane na powierzchni. Przedni mikrobiol (2019) 10:1691. doi: 10.3389/fmicb.2019.01691
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
208. Lieberman NAP, Lin MJ, Xie HI, Shrestha L, Nguyen T, Huang ML i in. Sekwencjonowanie genomu Treponema pallidum z sześciu kontynentów ujawnia zmienność genów kandydatów na szczepionkę i dominację szczepów kladu Nicols na Madagaskarze. PloS Negl Trop Dis (2021) 15(12):e0010063. doi: 10.1371/journal.pntd.0010063
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
209. Ovcinnikov NM, Delektorskij VV. Treponema pallidum we włóknach nerwowych. Br J. Venereal Dis (1975) 51(1):10. doi: 10.1136/sti.51.1.10
210. Sell S, Salman J. Wykazanie treponema pallidum w aksonach nerwów skórnych w doświadczalnych wrzodach królików. Sex Transm Dis (1992) 19(1):1–6. doi: 10.1097/00007435-199201000-00001
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
211. MA Medici. Nisza immunoprotekcyjna – nowy mechanizm patogenetyczny kiły, grzybic układowych i innych chorób zakaźnych. J. Theor Biol (1972) 36(3):617–25. doi: 10.1016/0022-5193(72)90012-4
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
212. Kronvall G, Simmons A, Myhre EB, Jonsson S. Specyficzna absorpcja albuminy surowicy ludzkiej, immunoglobuliny a i immunoglobuliny G z wybranymi szczepami paciorkowców grupy a i G. Infect Immun (1979) 25(1):1–10. doi: 10.1128/iai.25.1.1-10.1979
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
213. Alderete JF, Baseman JB. Związane z powierzchnią białka gospodarza na zjadliwym treponema pallidum. Infect Immun (1979) 26(3):1048–56. doi: 10.1128/iai.26.3.1048-1056.1979
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
214. Egesten A, Frick IM, Mörgelin M, Olin AI, Björck L. Wiązanie albuminy sprzyja przetrwaniu bakterii na powierzchni nabłonka. J Biol Chem (2011) 286(4):2469. doi: 10.1074/jbc.m110.148171
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
215. Champion CI, Miller JN, Borenstein LA, Lovett MA, Blanco DR. Szczepienie treponema pallidum endoflagella zmienia przebieg doświadczalnej kiły królików. Infect Immun (1990) 58(9):3158–61. doi: 10.1128/iai.58.9.3158-3161.1990
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
216. Zheng K, Xu M, Xiao Y, Luo H, Xie Y, Yu J i in. Immunogenność i skuteczność ochronna przeciwko treponema pallidum u królików nowozelandzkich immunizowanych plazmidowym DNA kodującym flagelinę. Emerg Microbes Infect (2018) 7(1):177. doi: 10.1038/s41426-018-0176-0
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
217. Nimmerjahn F., Ravetch JV. Receptory Fcγ jako regulatory odpowiedzi immunologicznej. Nat Rev Immunol (2008) 8(1):34–47. doi: 10.1038/nri2206
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
218. Lukehart SA, Molini B, Gomez A, Godornes C, Hof R, Fernandez MC i in. Szczepienie szczepionką przeciw kile składającą się z trzech antygenów znacząco osłabia rozwój wrzodu, zmniejsza obciążenie bakteryjne i hamuje rozprzestrzenianie się krętka bladego. Szczepionka (2022) 40(52):7676–92. doi: 10.1016/J.vaccine.2022.11.002
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
219. Haake DA, Mazel MK, McCoy AM, Milward F, Chao G, Matsunaga J i in. Białka błony zewnętrznej Leptospira OmpL1 i LipL41 wykazują synergistyczną immunoprotekcję. Infect Immun (1999) 67(12):6572–82. doi: 10.1128/iai.67.12.6572-6582.1999
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
220. Edmondson DG, Norris SJ. Hodowla in vitro krętka kiły treponema pallidum. Curr Protoc (2021) 1(2):e44. doi: 10.1002/cpz1.44
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
221. Phan A, Romeis E, Tantalo L, Giacani L. Transformacja in vitro i selekcja treponema pallidum subsp. blady. Curr Protoc (2022) 2(8):e507. doi: 10.1002/cpz1.507
Streszczenie PubMedu | Pełny tekst CrossRef | Google Scholar
- 1 Instytut Badań nad AIDS IrsiCaixa, Badalona, Hiszpania
- 2 Uniwersytet Autonomiczny w Barcelonie, Cerdanyola del Vallès, Katalonia, Hiszpania
- 3 Oddział Zaniedbanych Chorób Tropikalnych i Zakażeń Przenoszonych drogą płciową, Niemcy Szpital Trias i Pujol, Badalona, Hiszpania
- 4 Fundacja Fight Infections, Germans Trias i Pujol Hospital, Badalona, Katalonia, Hiszpania
- 5 Centrum Badań nad Zdrowiem i Opieką Społeczną (CESS), Wydział Lekarski, Uniwersytet Vic – Central University of Catalonia (UVic – UCC), Vic, Katalonia, Hiszpania
- 6 Germans Trias i Pujol Research Institute (IGTP), Badalona, Hiszpania
- 7 CIBERINFEC, Instituto de Salut Carlos III (ISCIII), Madryt, Hiszpania
Słowa kluczowe: kiła, krętek blady, szczepionka, odpowiedź immunologiczna, białka błony zewnętrznej
Cytat: Ávila-Nieto C, Pedreño-López N, Mitjà O, Clotet B, Blanco J i Carrillo J (2023) Szczepionka na kiłę: wyzwania, kontrowersje i możliwości. Przód. Immunol. 14:1126170. doi: 10.3389/fimmu.2023.1126170
Otrzymano: 17 grudnia 2022 r.; Zaakceptowano: 27 marca 2023 r.;
Opublikowano: 06 kwietnia 2023.
Edytowany przez:
Catherine Mary O’Connell z Uniwersytetu Karoliny Północnej w Chapel Hill, Stany Zjednoczone
Zrecenzowany przez:
Elizabeth De Gaspari , Instytut Adolfo Lutza, Brazylia
Kelly Lynn Hawley , Centrum Zdrowia Uniwersytetu Connecticut, Stany Zjednoczone
Immunol., 06 kwietnia 2023 r.
Rozdz. Szczepionki i terapie molekularne
Tom 14 – 2023 | https://doi.org/10.3389/fimmu.2023.1126170
Prawa autorskie © 2023 Ávila-Nieto, Pedreño-López, Mitjà, Clotet, Blanco i Carrillo. To jest artykuł o otwartym dostępie, rozpowszechniany na warunkach licencji Creative Commons Uznanie autorstwa (CC BY) . Wykorzystywanie, rozpowszechnianie lub powielanie na innych forach jest dozwolone pod warunkiem podania oryginalnego autora(ów) i właściciela(ów) praw autorskich oraz zacytowania oryginalnej publikacji w tym czasopiśmie, zgodnie z przyjętą praktyką akademicką. Zabronione jest wykorzystywanie, dystrybucja lub reprodukcja niezgodna z niniejszymi warunkami.
*Korespondencja: Jorge Carrillo, jcarrillo@irsicaixa.es ; Carlos Ávila-Nieto, cavila@irsicaixa.es
Zastrzeżenie: Wszystkie roszczenia wyrażone w tym artykule są wyłącznie roszczeniami autorów i niekoniecznie reprezentują roszczenia organizacji z nimi stowarzyszonych lub wydawcy, redaktorów i recenzentów. Żaden produkt, który może być oceniany w tym artykule lub twierdzenie jego producenta, nie jest objęte gwarancją ani poparciem wydawcy.
Link do artykułu: https://www.frontiersin.org/articles/10.3389/fimmu.2023.1126170/full