


Rolnictwo kukurydzy Azteków, ukazane w Kodeksie Florenckim. By Gary Francisco Keller, artwork created under supervision of Bernardino de Sahagún between 1540 and 1585. – The Digital Edition of the Florentine Codex, CC BY 3.0, https://commons.wikimedia.org/w/index.php?curid=15124157
Ilustrację tę łączy z poniższym artykułem tylko uprawa kukurydzy, oraz w jakimś sensie też okres, choć państwowość Azteków rozwijała się dopiero od XII wieku, a ilustracja w Kodeksie odnosi się do jej schyłku w XVI wieku. Warto jednak zwrócić uwagę na bardzo prymitywny sposób uprawy kukurydzy i proste narzędzia. Z jakiego powodu postęp w tej dziedzinie był tak niewielki? Pojawia się też pytanie, z jakiego powodu tak trudno znaleźć wśród aż tylu artefaktów kultury Majów takie, które przedstawiałyby pracę rolnika? Tym zajmowała się ludność poddana. Podobne zjawisko obserwować możemy w historii Egiptu i wielu innych starożytnych państwowości, dla których przyjęliśmy określenie „cywilizacji”. Zdobycze tej w państwach Majów i Azteków były podobne. W dziedzinie rolnictwa prawie wszędzie naprawdę niewielkie, wzbudzało ono niewielkie zainteresowanie władców.
Poniższy artykuł publikujemy z kilku powodów. Po pierwsze opisane w nim badania ukazują związek między zmianami klimatycznymi a rozwojem cywilizacji. Badania te, obejmujące aż tak długi okres, dowodzą ponadto, że zmiany klimatyczne i tzw. okresy ocieplenia występowały w stosunkowo bliskiej historii dość często. Oczywiście to nie działalność człowieka była ich przyczyną.
Odnośnie działalności człowieka warto postawić jeszcze jedno pytanie. Zazwyczaj, bowiem widząc tzw. osiągnięcia kultur Mezoameryki ogarnia nas zachwyt. Nad czym, pytam? Piramidami, które są świadectwem marnowania ludzkiej aktywności w celu tylko legitymizacji władzy politycznych przywódców, ich celów propagandowych. Z naszego punktu widzenia ich funkcjonalność była żadna. Cytowani poniżej badacze próbują z mozołem, dzięki najnowszym metodom badawczym, odtworzyć pewne fakty z życia ludów Mezoameryki w okresie aż 3 000 lat (mniej więcej od 1 500 roku p.n.e. do 1 5000 roku n.e. W pewnych aspektach cytowane badania odnoszą się do okresu jeszcze dłuższego, bo aż od 3000 lat p.n.e.). Jak niewielki był dorobek cywilizacyjny tych społeczeństw w tak długim okresie? Wbrew powszechnej opinii, wspomniany przecież przez cytowanych badaczy, dorobek w matematyce i astronomii był też, z dzisiejszego punktu widzenia, bardzo niewielki. Dlaczego? Przyczynami kolejnych regresów w dziejach społeczności Majów były zmiany klimatyczne, w tym wypadku okresy suszy. Czy jednak tylko one? A może to wojny i podboje oraz błędny model organizacji społeczeństw, na których kształtowanie te pierwsze tak długo wywierały zasadniczy wpływ, powstrzymywały ten postęp?
Warto poznawać historię, warto o niej czasami rozmyślać.
Redakcja
Susze, uprawy kukurydzy i rozwój państwowości starożytnych Majów
Raporty naukowe tom 12 , Numer artykułu: 22272 ( 2022 )
Abstrakt
Starożytna kultura Majów w Mezoameryce przez wieki kształtowała krajobrazy na obszarze, gdzie uprawiano kukurydzę ( Zea mays). Uprawa ta jest uważana za podstawową w diecie obecnych i starożytnych kultur mezoamerykańskich. Rejestracje pyłków z miejsc o różnych warunkach środowiskowych i klimatycznych na Półwyspie Jukatan (Meksyk) i Peten (Gwatemala) wskazują na wyraźny związek między zwiększonym stężeniem pyłku kukurydzy a okresami zmniejszonych opadów spowodowanych oscylacją południową El Niño (ENSO), natomiast okresy wilgotne charakteryzują się niską zawartością pyłku kukurydzy. Warunki ENSO nie były równomiernie rozłożone na półwyspie Jukatan, a regionalne susze różnią się w zależności od regionalnego klimatu i warunków geograficznych. Nasze wyniki wskazują na silny związek zwiększonego spadku upraw kukurydzy i lasów tropikalnych z okresami suszy, podczas gdy późny przedklasyczny okres wilgotny (ok. 500–200 pne) charakteryzuje się brakiem pyłku kukurydzy. Suchy późny preklasyk (300 pne-250 n.e.) był kluczowym okresem dla zwiększonej produkcji kukurydzy, co sugeruje nowe podejście do uprawy kukurydzy. Rola kukurydzy uległa zmianie, z podstawowej uprawy dietetycznej, do pragmatycznego produktu, który miał sprostać niekorzystnym warunkom środowiskowym
Wprowadzenie
Rozkwit starożytnej kultury Majów w Mezoameryce w ciągu prawie 3000 lat jest tradycyjnie związany ze skutecznymi praktykami rolniczymi, rozwojem kultury oraz postępem naukowym w matematyce i astronomii1,2. Rozwój kulturowy starożytnych Majów można podzielić na cztery okresy: wczesny, środkowy i późny preklasyczny (1500 pne do 250 n.e.) oraz wczesny do późnego klasycznego (250 do 1200 n.e.) 3 .
Między 900 a 1100 rokiem n.e. Majowie doświadczyli społecznych i politycznych przemian w warunkach suszy, co skomplikowało pełne zrozumienie przyczyn i procesu ich upadku4,5. Aby od okresu przedklasycznego mogło dochodzić do stałego, równomiernego rozwoju społecznego konieczne były wystarczające i ciągłe dostawy żywności. Ze względu na konieczność stałego zaopatrzenia w żywność za najważniejszą uprawę starożytnej kultury Majów uważa się kukurydzę2. Wiedza rolnicza Majów została opisana jako spuścizna pradawnych praktyk rolniczych 6, przy czym zidentyfikowano ponad 600 gatunków roślin o określonych zastosowaniach 7, 8 .
Istnieje coraz większa wiedza o tym, że starożytni Majowie mogli zarządzać krajobrazem jako ogrodem leśnym na skalę lokalną i regionalną 9 , 10 , 11. Beach i wsp.12 nazwali okres od 3000 do 1000 lat kal. BP (1050 p.n.e.– 950 n.e.) Majaceńskim, wskazując, że to wówczas starożytni Majowie przekształcili krajobraz i system hydrologiczny na szerszą skalę regionalną. Zmiana ta obejmowała przekształcenie terenów podmokłych i dostosowanie ich do użytku rolniczego. Pohl i in. 13 donosi o wykorzystaniu kukurydzy z terenów podmokłych północnego Belize około 3000 r. p.n.e., a od 2500 p.n.e. zintensyfikowaniu uprawy kukurydzy. Na północny Półwyspu Jukatan, w regionie Yalahau, tereny podmokłe były zarządzane za pomocą wałów i tam14 .
Przedmiotem analizy jest wpływ zmian klimatu w przeszłości na starożytnych Majów, ich adaptację kulturową i odporność środowiskową, ponieważ większość badań paleoekologicznych koncentrowała się na suszach na obszarach zajmowanych przez Majów z późnego okresu klasycznego. Starożytna kultura Majów przystosowała się do zmieniających się warunków klimatycznych i do różnych warunków środowiskowych15. Wiele badań paleoklimatycznych ujawniło wpływ zmienności klimatu na kulturę i środowisko starożytnych Majów, a kilka innych badań sugeruje również, że okresy suszy odegrały główną rolę w ich rozwoju kulturowym i upadku16,17. Okresy wilgotne, z naprzemiennymi krótkimi fazami suszy i przedłużającymi się suszami, zostały zidentyfikowane w szerszym regionie Majów w ciągu ostatnich 4000 lat16,18, a były spowodowane głównie przesunięciami równoleżnikowymi międzytropikalnej strefy konwergencji5. W badaniach tych uznano również wpływ oscylacji południowej El Niño (ENSO) na dynamikę klimatu i ekosystemy 4 , 5 , 16 , 19 , 20 , 21. Wpływ obecnej i przeszłej ENSO na południowy Meksyk i Amerykę Środkową jest znany z kilku badań 22 , 23. Badanie geochemiczne przeprowadzone w Lago Puerto Arturo w północnej Gwatemali sugeruje, że suche warunki środowiskowe sprzyjały rolnictwu w późnym holocenie24. I odwrotnie, bardziej wilgotne warunki w okresie preklasycznym hamowały rozwój intensywnej uprawy kukurydzy do około 400 roku p.n.e.25.
Niedawne ENSO w 2016 r. spowodowało zmniejszenie opadów i warunki stresu fizjologicznego na drzewach na obszarze Półwyspu Jukatan (YP) wzdłuż gradientu opadów, przy czym gatunki drzew z najsuchszych miejsc były bardziej dotknięte w porównaniu z gatunkami drzew z najbardziej wilgotnych miejsc26.
Dowody na uprawę kukurydzy od wczesnego okresu preklasycznego są dostępne z kilku zapisów dotyczących pyłków25, dlatego pozostaje pytanie, w jaki sposób susze oddziaływały na roślinność i zmianę użytkowania gruntów w okresie przedklasycznym i klasycznym. Szczegółowe spojrzenie na dane paleoekologiczne od czasów wczesnego preklasyku może ułatwić nam zrozumienie znaczenia roli kukurydzy używanej przez starożytnych Majów w okresach zmniejszonych opadów na rozwój ich cywilizacji.
Stawiamy hipotezę, że w suchszych warunkach klimatycznych w zapisach pyłkowych wykrywa się zwiększoną obecność pyłku kukurydzy. Stąd w wilgotnych warunkach klimatycznych obecność pyłku kukurydzy będzie mniejsza. Rejestry okrzemek i pyłków z jeziora Tuspan25 i Rio Hondo 20 wskazują na przejście do wilgotnych warunków klimatycznych, które panowały między 1000 a 850 p.n.e. Rosenswiga i in. 27 uważają ten okres, około 1000 roku p.n.e., za kluczowy dla rozwoju kulturowego, kiedy kukurydza zyskała na znaczeniu dla starożytnych kultur w Mezoameryce. Zdaniem Rosenswiga i in.27nie należy tego wnioskować na podstawie zmian paleoekologicznych, około 1000 cal p.n.e., podczas gdy ta data wskazuje na kluczowy okres dla kukurydzy, po poprzednim ciągłym trendzie suszy między 4500 a 2800 cal rokiem BP (2550-850 p.n.e.) w Mezoameryce. Po 3500 roku p.n.e. (1550 p.n.e.) w kilku zapisach paleoekologicznych z Półwyspu Jukatan można zaobserwować otwarcie krajobrazu. Wylesianie spowodowane zmianą klimatu poprzedziło produkcję kukurydzy na obszarze Peten-Itza około 1000 cal p.n.e. 28, ale nie jest to rozstrzygające dla wszystkich obszarów Półwyspu Jukatan.
Wyniki
Od wczesnego do środkowego preklasyku (1500–600 p.n.e.)
W przypadku wczesnego i środkowego preklasyku, nasze północne i najbardziej suche miejsca badawcze (Ria Lagartos 1 i 2) wskazują na rosnącą obecność kukurydzy wraz z silniejszą aktywnością ENSO około 1300-1100 p.n.e., 1000-800 p.n.e., 750-600 p.n.e.29, 3. Susze z okresu środkowego preklasyku wykazują różnice regionalne w obrębie obszaru dystrybucji starożytnych Majów. Wczesne i środkowe przejście przedklasyczne wykazało zmienność geograficzną, pokazując, że najbardziej wilgotne miejsce (Peten-Itza) wykazuje ujemną korelację Pearsona wynoszącą Peten-Itza r (30) = −0, 48, p = 0, 01 między pyłkiem lasów tropikalnych a ENSO. Suche warunki klimatyczne na wschodnim półwyspie Jukatan wykryto między 900 a 600 rokiem p.n.e. oraz na innych obszarach nizin Majów20, które mieszczą się w okresie 2800 cal BP31. Rdzeń Silvituc wykazuje znaczący pozytywny związek między lasem tropikalnym (r = 0,57), roślinnością zakłócającą (r = 0,59), kukurydzą (r = 0,63) i ENSO i mieści się w ramach czasowych między 800 a 600 p.n.e. osadnictwa w tym czasie okresu suszy. Stanowiska Ria Lagartos wykazały ujemną korelację między pyłkiem kukurydzy/tropikalnym a ENSO (Tabela 1 ).
Przejście od średniego do późnego preklasycznego (600 p.n.e. – 250 n.e.)
Między 550 a 200 rokiem p.n.e. odnotowano wilgotny okres, znany jako późny przedklasyczny okres wilgotny (LPHP)18, który zidentyfikowano w naszych rdzeniach osadów jako okres z niewielką ilością pyłku kukurydzy i zwiększonymi wartościami pyłku sklasyfikowanego jako elementy lasów tropikalnych. Szacowane średnie opady były do 20% wyższe podczas LPHP w porównaniu z wartościami obecnymi we wschodnim YP32. Współczynnik korelacji Pearsona obliczono dla zależności między taksonami lasów tropikalnych a ENSO. Dla tego podanego okresu nie wykryto żadnej korelacji, czego oczekiwano, biorąc pod uwagę warunki zwiększonego poziomu opadów.
Po LPHP obliczone współczynniki korelacji Pearsona wykazały korelację ENSO z mokrymi stanowiskami południowo-wschodniego półwyspu Jukatan, Rio Hondo i Silvituc (Rio Hondo r(3) = 0,84, p = 1; Silvituc r(22) = 0,37, p = 1; odpowiednio). Północne suche miejsca nie wykazały żadnej korelacji, podobnie jak bardziej wilgotne miejsce Peten-Itza. Obecność kukurydzy zaczęła wzrastać w naszych zapisach pyłkowych po redukcji lasów spowodowanej klimatem w Ria Lagartos, Chumpich, Peten-Itza, Silvituc i Rio Hondo w późnym preklasyku około 250-1 pne. Pyłek taksonów lasów tropikalnych rdzenia Rio Hondo silnie korelował z ENSO r(3) = 0,9, p = 1. Jesteśmy jednak świadomi małej liczby punktów danych dla tego rdzenia w tym przedziale czasowym.
Wczesny klasyk (250–500 n.e.)
We wczesnym okresie klasycznym elementy lasów tropikalnych na wszystkich stanowiskach wykazywały ujemną korelację z ENSO, z wyjątkiem jeziora Silvituc. Te ramy czasowe dobrze pasują do pierwszego opuszczenia obszaru Silvituc podczas wczesnego okresu klasycznego i zwiększonej obecności pyłku z roślinności wtórnej (Torrescano-Valle & Islebe, 2015), jak również uogólnionej redukcji pyłku kukurydzy we wszystkich innych zapisach pyłkowych.
Późny klasyk do klasycznego postu (500–1200 n.e.)
W okresie późnego klasyku północne suche stanowisko wykazuje dodatnią korelację między kukurydzą a ENSO (Ria Lagartos 1 r(15) = 0,46, p = 0,6). To dobrze pasuje do zwiększonej suszy zgłaszanej z północnego półwyspu Jukatan podczas późnego klasyku. Inne miejsca zgłoszone w tym badaniu wykazują ujemną korelację kukurydza / ENSO, ponieważ obszary te zostały opuszczone po 800 roku n.e.
Dyskusja
Wzrost obecności kukurydzy we wczesnym i środkowym okresie preklasycznym obserwuje się po rosnącym trendzie suszy przed 3500 rokiem kal. BP (1550 p.n.e.). Kukurydza zwiększyła swoją obecność w północnej Mezoameryce po roku 1000 p.n.e.27, co oznacza zwiększoną zmianę użytkowania gruntów w pobliżu zbiorników wodnych w tym regionie (ryc. 3 ). Wylesianie wywołane klimatem jest widoczne w 3000 roku kal. BP w jeziorach Peten-Itza 28 i jeziorze Silvituc 33, gdzie otwarciu krajobrazu sprzyjały zmniejszone opady.
Późny preklasyczny okres wilgotny (LPHP), z brakiem kukurydzy w zapisach pyłkowych, pośrednio wskazuje, że dostępne były inne zasoby roślin jadalnych. W tym okresie również w wielu zapisach paleoekologicznych nizin Majów 20 , 24 , 25 , 33 występuje wzrost wartości taksonów pyłków lasów tropikalnych. Po LPHP nastąpił wzrost kukurydzy we wszystkich zapisach, zwłaszcza podczas późnej suszy preklasycznej (150–250 n.e.), kiedy kukurydza znacznie zwiększyła swoją obecność w zapisach pyłkowych Ria Lagartos, Peten-Itza, Silvituc i Chumpich. Następnie obecność kukurydzy ponownie spadła po 250 roku n.e., gdy regionalne opady zaczęły się stabilizować 32. Późna susza przedklasyczna wykazała spadek opadów o co najmniej 20% w większości części YP w porównaniu z poprzednim okresem i była najsilniejszą suszą ostatnich 2000 lat na północnym półwyspie Jukatan. Zostało to również zaobserwowane w aktualnych danych ENSO 2016., które spowodowały spadek opadów nawet o 34% na północy Półwyspu Jukatan 26 . Jezioro Peten Itza jest najbardziej wilgotnym ze wszystkich miejsc, ze średnimi rocznymi opadami wynoszącymi ok. 1600 mm, zmniejszone opady mogły spowodować mniejszy stres (straty?) w lasach tropikalnych. W okresie preklasycznym pojawiła się bardziej zintensyfikowana forma rolnictwa będąca wynikiem adaptacja do zwiększonej liczby ludności i obniżonych poziomów opadów34,35. Zintensyfikowaną działalność rolniczą osiągnięto między innymi dzięki tarasowaniu, uprawie pól, nawadnianiu i kanałom35. Kukurydza mogłaby zatem być wykorzystywana z większą intensywnością rolnictwa w odpowiedzi na regionalne susze i wylesianie spowodowane klimatem lub zmianę użytkowania gruntów przez człowieka. Poza obszarem rdzenia starożytnych Majów, z Kostaryki, Kerr 36 przedstawia podobne dowody na wzrost upraw kukurydzy w okresach suszy. Uprawa kukurydzy w Laguna Bonillita osiągnęła szczyt w 150 roku n.e. podczas późnej suszy przedklasycznej.
Po suszy w późnym okresie przedklasycznym średni wzrost opadów ponownie odpowiada spadkowi procentowej zawartości pyłku kukurydzy we wszystkich rdzeniach (ryc. 3 ). Po 250 roku n.e. w społeczeństwach Majów zaobserwowano złożone procesy zmian społecznych wraz ze wzrostem liczby ludności na nizinach Majów Środkowych, aż do Terminalu Classic 37 .
Widma pyłkowe z okresu klasycznego świadczą o zmniejszonej obecności pyłku kukurydzy po ok. 300 n.e., dostarczając pośrednich dowodów na to, że główne praktyki agroleśne nie były uzależnione głównie od kukurydzy. Taksony roślinności zakłócającej są stale obecne w okresie klasycznym, spadając po okresie klasycznym terminalnym (ryc. 3 ). Susze dotknęły selektywnie półwysep Jukatan, biorąc pod uwagę geograficzne rozmieszczenie badanych rekordów pyłkowych. Czas, rozmieszczenie geograficzne i reakcja na czynniki środowiskowe wskazują na mozaikę zmian paleoekologicznych i paleoklimatologicznych. Dzisiejsza analiza anomalii klimatycznych Półwyspu Jukatan pokazuje, że istnieje pięć różnych skupisk anomalii opadów 38, efekt przestrzenny tych anomalii na roślinność jest szczególnie widoczny w warunkach ENSO przy zmniejszonych opadach26. Wszystkie sześć rekordów pyłkowych mieści się w tych skupiskach anomalii opadów. Widma pyłków kopalnych odzwierciedlają bardziej suche warunki środowiskowe w danych okresach i zostały wykryte w skali regionalnej30,39. Obecnie południowo-wschodnia i południowo-zachodnia część Półwyspu Jukatan są najbardziej wilgotnymi obszarami tego regionu, a kukurydza skorzystała na zmniejszonych opadach w tych regionach. Susza z późnego okresu preklasycznego miała najsilniejszy wpływ w połączeniu ze zmianami użytkowania gruntów przez człowieka na roślinność w południowo-wschodniej i południowo-zachodniej części (rdzenie Rio Hondo i Silvituc).
Dane kopalnych pyłków kukurydzy i ich związek ze zmiennymi poziomami opadów wskazują na zwiększoną produkcję kukurydzy po 300–200 p.n.e. w odpowiedzi na regionalne suche warunki klimatyczne, lokalne warunki środowiskowe i zaburzenia lasów tropikalnych. Starożytna kultura Majów nie opierała się wyłącznie na kukurydzy, ponieważ dostępnych było wiele innych gatunków roślin jadalnych. Zgadzamy się z Tuxillem i in.40, że kukurydza pomagała powstrzymać głód w okresach suszy. Geograficzny zasięg susz i ich wpływ na lasy tropikalne oraz uprawę kukurydzy jest kluczowym czynnikiem pozwalającym zrozumieć, jak rozwinęła się starożytna kultura Majów.
Metody
W tym badaniu przeanalizowaliśmy obecność skamieniałego pyłku kukurydzy z kilku rdzeni osadów Półwyspu Jukatan (YP) i północnej Gwatemali, związanych ze zmiennością ENSO w ciągu ostatnich ~ 4000 lat (ryc. 1 ). Analiza danych obejmuje numeryczną zależność wcześniejszych sygnałów ENSO w porównaniu z obecnością skamieniałego pyłku kukurydzy. Przeanalizowano sześć rdzeni osadów zlokalizowanych wzdłuż gradientu opadów (800–1500 mm/rok) w suchych obszarach lasów tropikalnych na Półwyspie Jukatan i w północnej Gwatemali. Chronologiczną kontrolę wszystkich rdzeni osiągnięto za pomocą datowania radiowęglowego AMS i oryginalnych danych kalibracyjnych. Wszystkie metody zostały przeprowadzone zgodnie z odpowiednimi wytycznymi i przepisami instytucjonalnymi, krajowymi i międzynarodowymi. Zastosowano następujące rdzenie (patrz informacje uzupełniające dla rozszerzonych danych z sześciu zapisów pyłkowych):
- 1.Ria Lagartos 1, położony w północnej części YP. Na obszarze tym średnie roczne opady wynoszą 500 mm, a średnia roczna temperatura wynosi 26ºC. W pobliżu tego głównego obszaru znajdują się sezonowo zalewane suche lasy i lasy namorzynowe. W tym miejscu pobrano rdzeń o długości 1,90 m 29 .
- 2.Ria Lagartos 2, patrz Ria Lagartos 1 w celu uzyskania informacji o miejscu 30 .
- 3.Jezioro Silvituc znajduje się w południowej części YP w stanie Campeche. Średnie roczne opady na tym obszarze wynoszą 1300 mm/rok, przy średniej rocznej temperaturze 26ºC. Pobrano rdzeń osadu o średnicy 1,35 m. Obecnie dominującym typem roślinności jest sezonowy suchy las tropikalny 33 .
- 4.Rio Hondo znajduje się w południowo-wschodniej części YP w stanie Quintana Roo. Pobrany rdzeń osadu ma długość 8 m i znajduje się na obszarze o rocznych opadach 1300–1400 mm, o średniej rocznej temperaturze 26,5 ºC. Roślinność składa się z mieszanki namorzynów i niskich lasów tropikalnych 20 .
- 5.Jezioro Chumpich znajduje się w południowej części YP, najbliższe stanowisko archeologiczne odpowiada Uxul, które jest ~50 km od Calakmul i ~30 km od El Mirador. Jezioro ma 1,7 km długości i 1,3 km szerokości, otoczone jest niskim zalewowym lasem, który łączy je z innymi pobliskimi jeziorami, takimi jak San Felipe i dorzecze Mirador (Wahl i in., 2007). Opady wahają się od 1300 do 1500 mm rocznie. Roślinność składa się głównie z tropikalnych lasów średnich, charakteryzujących się obecnością Brosimum alicastrum , Manilkara zapota , Talisia olivaeformis i Pimenta dioica . Niski las prezentuje gatunki takie jak Haematoxylum campechianum, Metopium brownei, Bucida buceras, Cladium jamaicense i Cyperaceae 41 .
- 6.Jezioro Petén-Itza znajduje się w północnej Gwatemali, w dystrykcie Peten. Aby zapoznać się z charakterystyką jeziora, patrz Islebe i in. 17 . Rdzeń miał 5,45 m długości. Średnie roczne opady wynoszą około 1600 mm, a średnia roczna temperatura wynosi 25ºC. Roślinność wokół jeziora składa się głównie z lasów tropikalnych z gatunkami m.in. z rodzin Moraceae i Fabaceae.

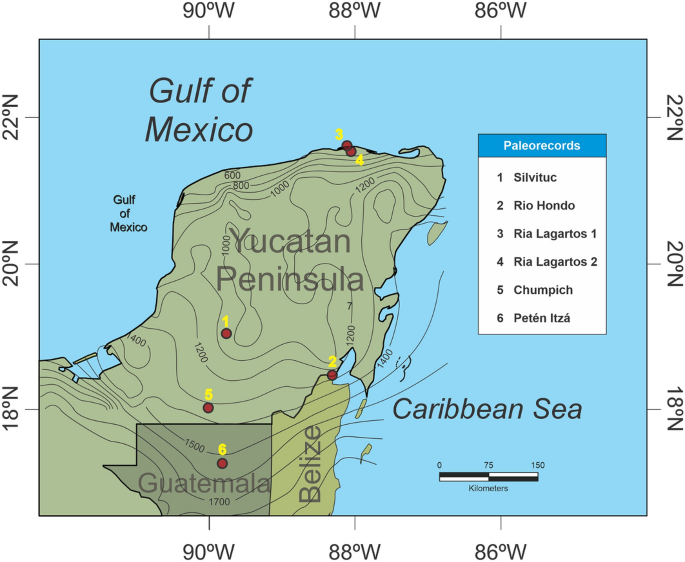
Główne lokalizacje na Półwyspie Jukatan i w północnej Gwatemali. Izohiety opadów pokazujące aktualne wartości opadów. Wygenerowane za pomocą QGIS wersja 3.22.11 58 ( https://qgis.org ) i izohiety zostały narysowane na podstawie mapy opadów z INEGI 59 .
W Tabeli 2 przedstawiono wszystkie taksony z sześciu zapisów pyłkowych.
Ekstrakcja pyłku
Do ekstrakcji pyłku ze wszystkich rdzeni osadów zastosowano acetolizę 42, a szczegółowe metody odsyłamy do oryginalnych publikacji. Oryginalne diagramy pyłkowe są dostępne w odpowiednich odnośnikach. Zidentyfikowane ziarna pyłku Zea mays wszystkich rdzeni mieściły się w zakresie od 65 do 90 mikronów. Dodatkowe szkiełka pyłkowe z różnych rdzeni skanowano pod kątem Z. mays , aby uniknąć brakujących ziaren. Zea mays zidentyfikowano na podstawie wielkości i morfologii porów, aw razie wątpliwości zastosowano kontrast fazowy.
Zea mays
Wśród głównych źródeł pożywienia wykorzystywanych przez kulturę Majów w okresie przedklasycznym i klasycznym były Brosimum alicastrum i Zea mays43,44. Drzewa B. alicastrum (orzechy chlebowe) są powszechne w okolicach obszarów milpa45, a ich nasiona są używane do dziś przez rolników Majów46. Brosimum alicastrum mogło być ważnym źródłem pożywienia dla starożytnych Majów. Gospodarowanie orzeszkami chlebowymi zostało potwierdzone przez analizę węgla drzewnego z północnej Gwatemali 47, która wskazuje na szerokie wykorzystanie tego gatunku drzew.
Majowie z Yucatec wybrali odmiany kukurydzy o różnych cyklach, a mianowicie kukurydzę o krótkim cyklu, trwającym od 7 do 10 tygodni, oraz kukurydzę o długim cyklu, trwającą 12 i 16 tygodni40. Czas trwania cyklu pozwala kukurydzy uniknąć wewnątrzsezonowej suszy, która występuje w porze deszczowej. Ponieważ sadzenie następuje na początku pory deszczowej, susza wewnątrzsezonowa może zbiegać się z dwoma kluczowymi fazami rozwojowymi i zwiększonym zapotrzebowaniem na wodę, takimi jak rozwój kłosów lub wzrost ziaren kukurydzy w kolbie 40. Z tego powodu Majowie tradycyjnie mieszali różne odmiany kukurydzy na tym samym poletku. Odmiany kukurydzy o krótkim cyklu zapewniają rozwój kłosów przed sezonową suszą i są znane jako nal t’eel, xt’uup nal i x-mejen nal. Odmiany o długim cyklu gwarantują wyższą produkcję ziarna w przypadku dobrej pory deszczowej i są znane jako X-nuuk nal, Ts’ı´it’it bakal, Bek’ech bakal 40. Oczekiwana produkcja tradycyjnej odmiany Maya milpa wynosi 2,5 tony ha-1 na cykl sadzenia 48. Podczas gdy produkcja B. alicastrum zależy od wieku, a drzewa w wieku 25 lat produkują do 19 ton rocznie świeżych nasion 49, co odpowiada 4 tonom suchych nasion rocznie o wilgotności do 80% 50. Jednak wpływ suszy i rosnących temperatur na produkcję biomasy zależy od stopnia przystosowania gatunków do stresu wodnego. Efektywność wykorzystania wody (WUE) wykazała związek między produktywnością roślin a zużyciem wody 51 . Z tego punktu widzenia kukurydza ma przewagę, ponieważ jej produktywność może wzrosnąć bez zmiany tempa zużycia wody, osiągając wzrost WUE 52 . Niektóre uprawy kukurydzy mogą odnotować WUE na poziomie 5 μmol mol -1 , podczas gdy B. alicastrum wykazuje maksymalną WUE na poziomie 1,5 μmol mol -1 49. Ponadto strategie przycinania mogą poprawić WUE. Zwiększenie liczby rzędów i gęstości nasadzeń pomaga zmniejszyć wpływ na parowanie wody z gleby i jest skuteczną strategią w regionach suchych ze zmiennymi epizodami opadów i dużym zapotrzebowaniem na parowanie 53 .
Analiza danych
Podejście interpolacyjne Zea Maysa
Liczebność pyłków na każdym stanowisku podzielono na typy roślinności: Zea mays , roślinność zakłócająca i lasy tropikalne (ryc. 2 ). Standaryzację danych przeprowadzono za pomocą funkcji Z-score, aby uzyskać porównywalne wartości między zapisami pyłków z miejsc Rı́o Hondo, Ria Lagartos, Silvituc, Chumpich, Peten Itza i zmienności ENSO w jeziorze Pallcacocha (intensywność koloru czerwonego) 54 . Aby ustalić zgodność czasową między witrynami, zastosowano oprogramowanie statystyczne R w wersji 4.0.5 (R Core Team, 2021), zastosowano funkcję interpolacji pakietu „psych” 55 wraz z „readxl” 56 i „corrplot” 57paczki. Zmienność ENSO dostarczyła modelowi chronologię, a 20-letni przedział czasowy został wykorzystany do modelowania interpolowanych wartości w każdym zapisie pyłkowym, zachowując niezmienioną pierwotną chronologię zapisów pyłkowych. Dane z Laguna Pallcacocha, których nie można było sparować, nie zostały wykorzystane. Interpolacji dokonano w celu uzyskania porównywalnych danych dotyczących pyłku we wszystkich miejscach iw tych samych ramach czasowych, bez utraty danych dotyczących pyłku. Obliczone dane wyjściowe porównano z każdą oryginalną krzywą danych, aby zweryfikować trend nowo modelowanych danych w stosunku do oryginalnych danych. Po uzyskaniu modelowanych danych pyłkowych z poszczególnych zapisów, wykreślono złożony zestaw danych Z wyników danych pyłkowych dla każdego typu roślinności na ryc. 3. Średnie wartości pyłku dla każdego typu roślinności były wizualnie porównywalne ze zmiennością ENSO. Dla każdego stanowiska uzyskano współczynniki korelacji Pearsona między typami roślinności a ENSO (Tabela 1 ). Analizie wskaźników korelacji dokonano według okresów okupacji Majów, od wczesnego preklasyku do późnego klasycyzmu. Ten przedział czasowy odpowiada w przybliżeniu ostatnim 3500 latom ( Informacje uzupełniające ).

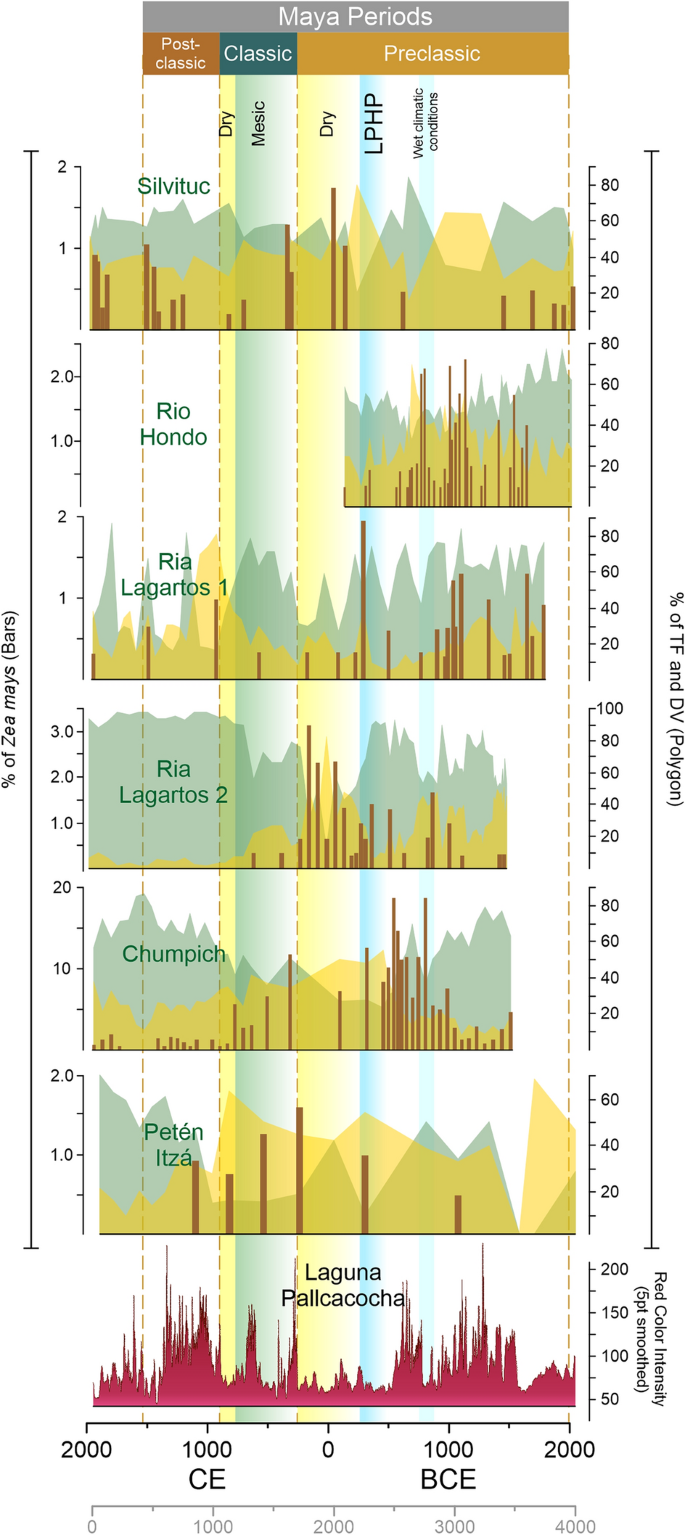
Diagramy pyłkowe rdzeni w obszarze Majów w YP. Procent lasów tropikalnych (TF; zacieniony na zielono) i roślinności zakłócającej (DV; zacieniony na żółto) wraz z Zea mays (brązowe słupki) i wygładzoną intensywnością czerwonego koloru w Laguna Pallcacocha 54 jako sygnał aktywności ENSO.

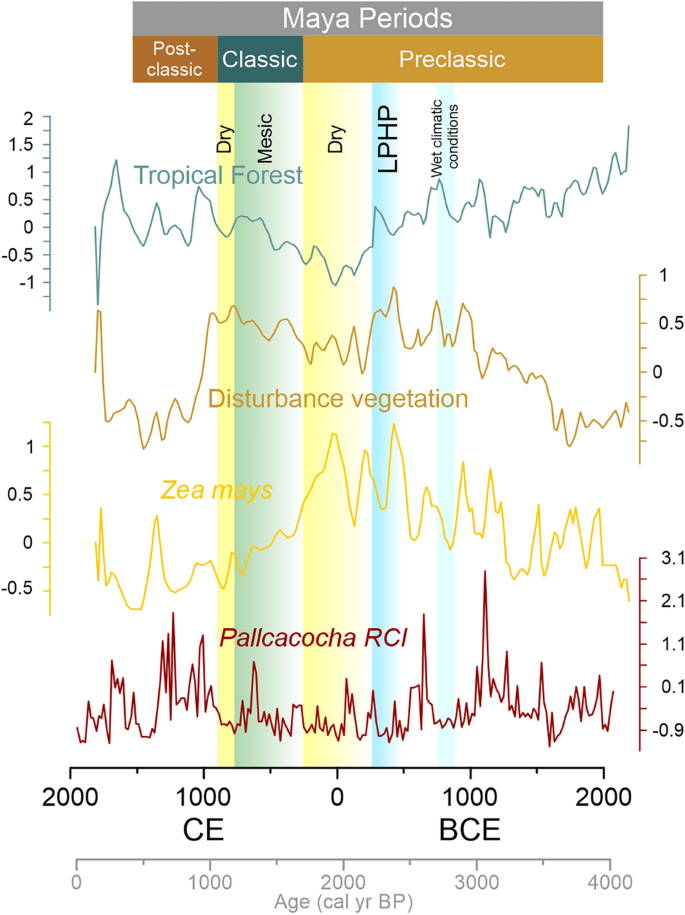
Średnie wartości Z-score grup pyłków kopalnych: lasy tropikalne, roślinność zakłócająca, Zea mays i Pallcacocha RCI (intensywność koloru czerwonego) jako zmienność ENSO 54 . W kolumnach wyróżniono okresy wcześniej zgłaszanej ogólnej zmienności klimatu.
Dostępność danych
Zbiory danych wykorzystane i/lub przeanalizowane podczas bieżącego badania są dostępne u odpowiedniego autora na uzasadnione żądanie.
Bibliografia
-
Evans, NP i in. Kwantyfikacja suszy podczas upadku klasycznej cywilizacji Majów. Nauka 361 , 498–501 (2018).
-
Gill, RB Wielkie susze Majów: woda, życie i śmierć (University of New Mexico Press, 2001).
-
Coe, MD Maya (Tamiza i Hudson, 1993).
-
Douglas, PMJ i in. Susza, adaptacja rolnictwa i upadek społeczno-polityczny na Nizinie Majów. proc. Natl. Acad. nauka USA 112 , 5607-5612 (2015).
-
Haug, GH i in. Klimat i upadek cywilizacji Majów. Nauka 299 , 1731-1735 (2003).
-
Ford, A. & Nigh, R. Pochodzenie leśnego ogrodu Majów: zarządzanie zasobami Majów. J. Etnobiol. 29 , 213-236 (2009).
-
Anderson, EN i in. Las Plantas de los Mayas: Etnobotánica en Quintana Roo, México (CONABIO-ECOSUR, 2005).
-
Fedick, SL Maya róg obfitości: rdzenne rośliny jadalne z nizin Majów. w The Real Business of Ancient Maya Economies (red. Masson, MA, Freidel, DA i Demarest, AA). 224–237 (University Press Floryda, 2020).
-
Ford, A. i Clarke, KC Łączenie przeszłości i teraźniejszości starożytnych Majów: użytkowanie gruntów na nizinach, rozmieszczenie ludności i gęstość w okresie późnego klasycznego. w The Oxford Handbook of Historical Ecology and Applied Archeology (red. Isendahl, C. & Stump, D.) (Oxford Handbook of Historical Ecology and Applied Archaeology, 2015).
-
Ford, A. & Nigh , R. Ogród leśny Majów : osiem tysiącleci zrównoważonej uprawy lasów tropikalnych , 2016).
-
Gómez-Pompa, A. O hodowli lasu Majów. meksykański ogier. (Estudios Mexicanos) 3 , 1–17 (1987).
-
Beach, T., Luzzadder-Beach, S., Krause, S. & Walling, S. Równina zalewowa i tereny podmokłe „Mayacene” w zlewni Rio Bravo w północno-zachodnim Belize. Holocen 25 (10), 1612–1622 (2015).
-
Pohl, MD i in. Wczesne rolnictwo na nizinach Majów. łac. Jestem. Antyk. 7 , 355-372 (1996).
-
Fedick, SL Zarządzana mozaika: rolnictwo starożytnych Majów i wykorzystanie zasobów (University of Utah Press, 1996).
-
Mueller, AD i in. Odbudowa ekosystemu leśnego na tropikalnych nizinach północnej Gwatemali po rozpadzie klasycznej polityki Majów. Geologia 38 , 523–526 (2010).
-
Hodell, DA, Curtis, JH & Brenner, M. Możliwa rola klimatu w upadku klasycznej cywilizacji Majów. Przyroda 375 , 391-394 (1995).
-
Islebe, GA, Hooghiemstra, H., Brenner, M., Curtis, JH & Hodell, DA Holoceńska historia roślinności z nizinnej Gwatemali. Holocen 6 , 265-271 (1996).
-
Medina-Elizalde, M., Polanco-Martínez, JM, Lases-Hernández, F., Bradley, R. & Burns, S. Testowanie hipotezy „burzy tropikalnej” dotyczącej zmienności klimatu Półwyspu Jukatan podczas klasycznego okresu Majów. Czwart. Rez. 86 , 111–119 (2016).
-
Aragón-Moreno, AA, Islebe, GA, Torrescano-Valle, N. & Arellano-Verdejo, J. Środkowa i późna holoceńska dynamika namorzynów na Półwyspie Jukatan w Meksyku. J. South Am. Nauka o Ziemi. 85 , 307–311 (2018).
-
Aragón-Moreno, AA, Islebe, GA, Roy, PD, Torrescano-Valle, N. i Mueller, AD Wymuszanie klimatu na roślinność południowo-wschodniego półwyspu Jukatan (Meksyk) w okresie od środkowego do późnego holocenu. Paleogeogr. Paleoklimatol. Paleoekol. 495 , 214–226 (2018).
-
Kennett, DJ i in. Rozwój i dezintegracja systemów politycznych Majów w odpowiedzi na zmiany klimatyczne. Nauka 338 , 788–791 (2012).
-
Conde, C. i in. El Niño i rolnictwo. w Los impactos de El Niño en México (red. Magaña, V.). 103–135 (Dirección General de Protección Civil, Secretaría de Gobernación, México, 1999).
-
Magaña, VO, Vázquez, JL, Pérez, JL & Pérez, JB Wpływ El Niño na opady atmosferyczne w Meksyku. Geofísica Int. 42 , 313-330 (2003).
-
Wahl, D., Byrne, R. i Anderson, L. Rekonstrukcja paleoklimatyczna sprzed 8700 lat z południowych nizin Majów. Czwart. nauka Obj. 103 , 19–25 (2014).
-
Nooren, K. i in. Wpływ klimatu na rozwój przedklasycznej cywilizacji Majów. wspiąć się Przeszłość 14 , 1253–1273 (2018).
-
Palomo-Kumul, J., Valdez-Hernández, M., Islebe, GA, Cach-Pérez, MJ i El Andrade, JL Oscylacja Niño-Southern wpływa na stosunki wodne gatunków drzew na Półwyspie Jukatan. Meksyk. nauka Rep. 11 , 10451 (2021).
-
Rosenswig, RM, VanDerwarker, AM, Culleton, BJ & Kennett, DJ Czy to już rolnictwo? Zintensyfikowane wykorzystanie kukurydzy w 1000 cal pne w Soconusco i Mezoameryce. J. Antropol. Archeol. 40 , 89-108 (2015).
-
Mueller, AD i in. Suszenie klimatu i związane z nim zanikanie lasów na nizinach północnej Gwatemali w późnym holocenie. Czwart. Rez. 71 , 133-141 (2009).
-
Aragón-Moreno, AA, Islebe, GA i Torrescano-Valle, N. A ~ 3800 lat, zapis w wysokiej rozdzielczości roślinności i zmian klimatu na północnym wybrzeżu Półwyspu Jukatan. Wielebny Paleobot. Palynol. 178 , 35–42 (2012).
-
Carrillo-Bastos, A., Islebe, GA i Torrescano-Valle, N. 3800 lat ilościowej rekonstrukcji opadów z północno-zachodniego półwyspu Jukatan. PLoS JEDEN 8 , e84333 (2013).
-
Berglund, BE Wpływ człowieka i zmiany klimatu — zdarzenia synchroniczne i związek przyczynowy?. Czwart. Int. 105 , 7–12 (2003).
-
Vela-Peláez, AA, Torrescano-Valle, N., Islebe, GA, Mas, JF i Weissenberger, H. Holoceńskie zmiany opadów w lesie Majów na półwyspie Jukatan. Meksyk. Paleogeogr. Paleoklimatol. Paleoekol. 505 , 42–52 (2018).
-
Torrescano-Valle, N. & Islebe, GA Paleoekologia holocenu, historia klimatu i wpływ człowieka na południowo-zachodnim półwyspie Jukatan. Wielebny Paleobot. Palynol. 217 , 1–8 (2015).
-
Anselmetti, FS, Hodell, DA, Ariztegui, D., Brenner, M. & Rosenmeier, MF Kwantyfikacja wskaźników erozji gleby związanych z wylesianiem starożytnych Majów. Geologia 35 , 915–918 (2007).
-
Plaża, T. i in. Przegląd zmian ludzkich i naturalnych na terenach podmokłych Niziny Majów w holocenie. Czwart. nauka Obj. 28 , 1710–1724 (2009).
-
Kerr, MT Zmienność opadów w holocenie, rolnictwo prehistoryczne oraz pożary naturalne i wywołane przez człowieka w Kostaryce (University of Tennessee, 2019).
-
Ebert, CE, Peniche May, N., Culleton, BJ, Awe, JJ & Kennett, DJ Regionalna odpowiedź na suszę podczas formowania się i upadku społeczeństw Majów przedklasycznych. Czwart. nauka Obj. 173 , 211–235 (2017).
-
De la Barreda, B., Metcalfe, SE & Boyd, DS Regionalizacja opadów, anomalie i występowanie suszy na Półwyspie Jukatan w Meksyku. Int. J. Climatol. 40 , 4541–4555 (2020).
-
Islebe, GA i in. Holoceńska paleoekologia i paleoklimatologia południowego i południowo-wschodniego Meksyku: podejście palinologiczne. w Meksykańskiej historii holocenu i antropocenu środowiska ( red. Torrescano-Valle, N., Islebe, GA i Roy, P.) (Springer, 2019).
-
Tuxill, J., Reyes, LA, Moreno, LL, Uicab, VC i Jarvis, DI. Każda kukurydza nie jest równa: wybór odmian kukurydzy i potrawy Majów na wiejskim Jukatanie w Meksyku. w Pre-Columbian Foodways: interdyscyplinarne podejście do żywności, kultury i rynków w starożytnej Mezoameryce (red. Staller, J. & Carrasco, M.) 467–486 (Springer, 2010).
-
Torrescano-Valle, N., Ramírez-Barajas, PJ, Islebe, GA, Vela-Pelaez, AA i Folan, WJ Wpływ człowieka a naturalna zmienność klimatu. w The Holocene and Anthropocene Environmental History of Mexico: A Paleoecological Approach on Mesoamerica (red. Torrescano-Valle, N., Islebe, GA & Roy, PD). 171–194 (Springer, 2019).
-
Faegri, K. & Iversen, J. Podręcznik analizy pyłków (Wiley, 1989).
-
Ford, A. Las Majów: udomowiony krajobraz. w The Maya World (red. Hutson, SR & Ardren, T.). 519–539 (Routledge, 2020).
-
Fedick, SL i Santiago, LS Duże zróżnicowanie dostępności źródeł roślin jadalnych Majów podczas starożytnych susz. proc. Natl. Acad. nauka Stany Zjednoczone 119 , 2115657118 (2022).
-
Puleston, DE Rola ramóna w utrzymaniu Majów. w Maya Subsistence . 353–366 (Elsevier, 1982).
-
Atran, S. i in. Tropikalne rolnictwo i leśnictwo Itza Maya [oraz komentarze i odpowiedzi]. bież. Antropol. 34 , 633-700 (1993).
-
Dussol, L., Elliott, M., Michelet, D. & Nondédéo, P. Starożytna hodowla Majów z orzecha zwyczajnego ( Brosimum alicastrum Sw.) i sapodilla ( Manilkara zapota (L.) P. Royen) w Naachtun (Gwatemala): A rekonstrukcja na podstawie analizy węgla drzewnego. Czwart. Int. 457 , 29–42 (2017).
-
Ebel, R., de Jesús Méndez Aguilar, M. i Putnam, HR Milpa: Jedna siostra zachorowała na klimat. Wpływ zmian klimatycznych na tradycyjne systemy rolnicze Majów. Int. J. Socjol. Rolnictwo. Jedzenie (online) 24 , 175–199 (2018).
-
Hernández-González, O. & Vergara-Yoisura, S. Badania nad produktywnością Brosimum alicastrum , drzewa tropikalnego wykorzystywanego jako pasza dla zwierząt na Półwyspie Jukatan. Bothalia 22 , 7 (2014).
-
Martínez-Ruiz, N. del R. i Larqué-Saavedra, A. Semilla de Ramón. w Alimentos Vegetales Autóctonos Iberoamericanos Subutilizados (red. Sonia, SA i Álvarez-Parrilla, E.). 177–192 (Fabro Editores, 2018).
-
Hatfield, JL & Dold, C. Efektywność zużycia wody: postępy i wyzwania w zmieniającym się klimacie. Z przodu. Nauka o roślinach. 10 , 103 (2019).
-
Basso, B. & Ritchie, JT Ewapotranspiracja w wysokowydajnej kukurydzy i przy zwiększonym deficycie prężności pary na Środkowym Zachodzie Stanów Zjednoczonych. Rolnictwo. Otaczać. Łotysz. 3 , 170039 (2018).
-
Gregory, PJ, Simmonds, LP & Pilbeam, CJ Rodzaj gleby, reżim klimatyczny i reakcja efektywności wykorzystania wody na zarządzanie uprawami. Agron J.92 , 814-820 (2000).
-
Moy, CM, Seltzer, GO, Rodbell, DT i Anderson, DM Zmienność aktywności El Niño/Southern Oscillation w tysiącletnich skalach czasowych w epoce holocenu. Przyroda 420 , 162–165 (2002).
-
Revelle, W. psych: Procedury badań psychologicznych, psychometrycznych i osobowości. Pakiet R na https://CRAN.R-project.org/package=psych (2022).
-
Wickham, H. & Bryan, J. readxl: Czytaj pliki Excela. Pakiet R pod adresem https://readxl.tidyverse.org/ (2022).
-
Wei, T. i in . Pakiet „corrplot”. Statystyk 56 , e24 (2017).
-
Zespół programistów QGIS. System informacji geograficznej QGIS. Stowarzyszenie QGIS na https://www.qgis.org (2022).
-
Instituto Nacional de Estadistica Geographia e Informatica (INEGI). 1:1000000 Merida, Carta de Precipitacion . Merida, Jukatan, Meksyk (1981).
Podziękowanie
Conacyt jest uznawany za finansowanie projektów Las selvas de la península de Yucatán durante el Holoceno Medio y Tardío: una compleja interacción de dinámica y resiliencia oraz Modelación Paleoclimática en la Península de Yucatán.
Deklaracje etyczne
Konkurujące interesy
Autorzy deklarują brak sprzecznych interesów.
Dodatkowe informacje
Uwaga wydawcy
Springer Nature pozostaje neutralny w odniesieniu do roszczeń jurysdykcyjnych na opublikowanych mapach i przynależności instytucjonalnej.
Link do artykułu:
https://www.nature.com/articles/s41598-022-26761-3
Prawa i uprawnienia
Otwarty dostęp Ten artykuł jest objęty licencją Creative Commons Attribution 4.0 International License, która zezwala na używanie, dzielenie się, adaptację, dystrybucję i powielanie na dowolnym nośniku lub w dowolnym formacie, pod warunkiem, że podasz odpowiednie oznaczenie oryginalnego autora (autorów) i źródła, podać link do licencji Creative Commons i wskazać, czy dokonano zmian. Obrazy lub inne materiały osób trzecich zawarte w tym artykule są objęte licencją Creative Commons, chyba że zaznaczono inaczej w informacji o autorze materiału. Jeśli materiał nie jest objęty licencją Creative Commons, a zamierzone użycie jest niezgodne z przepisami prawa lub wykracza poza dozwolone użycie, musisz uzyskać pozwolenie bezpośrednio od właściciela praw autorskich. Aby zobaczyć kopię tej licencji, odwiedźhttp://creativecommons.org/licenses/by/4.0/.
Techniki rolnicze Mezoameryki
Jednym z największych wyzwań dla rolników w Mezoameryce był brak gruntów użytkowych i zły stan gleby. Dwa główne sposoby zwalczania złej jakości gleby lub braku składników odżywczych w glebie to pozostawianie pól odłogiem przez pewien czas w cyklu milpa oraz stosowanie technik cięcia i wypalania. Znaczna część zapasów żywności Majów była uprawiana w ogrodach, znanych jako pet kot . System bierze swoją nazwę od niskiego muru z kamieni (pet oznacza okrągły i kot mur z luźnych kamieni), który charakterystycznie otacza leśną działkę ogrodową. Najwcześniejsze datowane kolby kukurydzy zostały odkryte w jaskini Guilá Naquitz w Oaxaca i pochodzą z 4300 pne. Kukurydza powstała w wyniku udomowienia teosinte , który jest uważany za przodka kukurydzy. Kukurydzę można przechowywać przez długi czas, można ją zmielić na mąkę i z łatwością zapewnia nadwyżki do wykorzystania w przyszłości. Kukurydza była niezbędna do przetrwania ludu Mezoameryki. Jego kulturowe znaczenie znajduje odzwierciedlenie w mitach, dziełach sztuki i rytuałach pochodzenia mezoamerykańskiego.
Rdzenni mieszkańcy Mezoameryki również stosowali techniki irygacyjne, podobne do innych wczesnych społeczności rolniczych we wczesnej Mezopotamii. Jednak w przeciwieństwie do suchych równin Żyznego Półksiężyca obszar Mezoameryki ma bardziej nierówny teren, przez co nawadnianie jest mniej skuteczne niż uprawy tarasowe i techniki cięcia i wypalania.
Techniki slash-and-burn to rodzaj ekstensywnego rolnictwa, w którym ilość pracy jest minimalna w pielęgnacji gruntów rolnych. Rolnictwo ekstensywne wymaga mniej siły roboczej, ale ma większy wpływ na otaczający je obszar. W przeciwieństwie do tego, rolnictwo intensywne odnosi się do rolnictwa, które wymaga dużych nakładów pracy, pozwalając na większe plony z tego samego obszaru, dzięki czemu lepiej nadaje się do siedzącego trybu życia.
Obraz wyróżniający: Rolnictwo kukurydzy Azteków, jak przedstawiono w Kodeksie Florenckim. By Gary Francisco Keller, artwork created under supervision of Bernardino de Sahagún between 1540 and 1585. – The Digital Edition of the Florentine Codex, CC BY 3.0, https://commons.wikimedia.org/w/index.php?curid=15124157