


Denisowa 4, ząb trzonowy. Z Wikipedii, wolnej encyklopedii.
Streszczenie
Obecność rodzaju Homo w plejstocenie w kontynentalnej Azji Południowo-Wschodniej jest przede wszystkim potwierdzona przez skąpe zapisy narzędzi kamiennych i rzadkie szczątki ludzkie. W niniejszym artykule przedstawiamy okaz hominina ze środkowego plejstocenu z Laosu, wraz z odkryciem zęba trzonowego z wapiennej jaskini Tam Ngu Hao 2 (Jaskinia Cobra) w Górach Annamickich. Wiek brekcji zawierającej skamieniałości waha się od 164 do 131 tys. lat, na podstawie bayesowskiego modelowania datowania luminescencyjnego matrycy osadowej, z której została wydobyta, datowania metodą serii U nadległego nacieku oraz datowania metodą serii U–ESR powiązanych zębów fauny. Analizy wewnętrznej struktury zęba trzonowego w połączeniu z paleoproteomicznymi analizami szkliwa wskazują, że ząb pochodzi od młodego, prawdopodobnie żeńskiego osobnika Homo . Bliskie podobieństwo morfologiczne do okazu Xiahe z Chin wskazuje, że należą one do tego samego taksonu i że Tam Ngu Hao 2 najprawdopodobniej jest przedstawicielem denisowianina.
Wstęp
Od wczesnego do późnego plejstocenu obecność Homo erectus jest dobrze udokumentowana w Azji, zwłaszcza w Chinach i Indonezji 1 , 2 , 3 . Jednak przypisanie taksonomiczne większości azjatyckich okazów Homo z późnego środkowego plejstocenu pozostaje kwestią sporną 4 , 5 , 6 , 7 . Niedawny opis i analiza czaszki z Harbin z Chin ponownie rozpaliły tę debatę, sugerując przypisanie jej nowemu gatunkowi o nazwie Homo longi 8 , ale ta nowa atrybucja taksonomiczna pozostaje przedmiotem gorących dyskusji. W rzeczywistości czaszka z Harbin wykazuje bliskie podobieństwo morfologiczne do innych azjatyckich okazów Homo z późnego środkowego do wczesnego późnego plejstocenu z Dali, Xujiayao, Xuchang i Hualongdong, których taksonomia pozostaje niejasna 4 , 9 , 10 . Uważa się, że te skamieniałości należą do innego taksonu niż H. erectus i często grupuje się je pod ogólną etykietą „archaiczni ludzie” 9 , 10 . Ze względu na kombinację cech, które wykazują, w tym cechy podobne do neandertalczyków, zasugerowano, że należą do azjatyckiego taksonu siostrzanego neandertalczyków, denisowian, nawet jeśli przypisanie to pozostaje przedmiotem dyskusji 5 , 11 , 12 . Niewielka liczba skamieniałości obecnie z całą pewnością przypisywanych denisowianom (Denisova 2, dolny lewy trzonowiec; Denisova 3, dystalny paliczek ręczny; Denisova 4, górny lewy M3; Denisova 8, górny trzonowiec; i żuchwa Xiahe) 13 , 14 , 15 , 16 uniemożliwia uzyskanie jasnego obrazu morfologicznego ich ogólnej morfologii. Rozmieszczenie geograficzne denisowian również pozostaje przedmiotem dyskusji. Współcześni Papuasi, Aborygeni australijscy, Oceania/Melanezyjczycy, filipińskie grupy Ayta i, w znacznie mniejszym stopniu, populacje kontynentalnej Azji Południowo-Wschodniej, zachowują dziedzictwo genetyczne Denisowian14 , 17 , 18 , 19. Połączone analizy paleoproteomiczne i morfometrycze niedawno zasugerowały, że żuchwa Xiahe ze środkowego plejstocenu z jaskini krasowej Baishiya należała do Denisowian, co rozszerza znany zasięg tej grupy na Wyżynę Tybetańską15 .. Nadal jednak nie ma dowodów kopalnych wyjaśniających odcisk genetyczny denisowian na współczesnych populacjach Azji Południowo-Wschodniej i — ze względu na niedobór zapisu kopalnego ze środkowego plejstocenu — nadal nie wiadomo, czy jedna lub więcej linii ludzkich (współ)istniało w kontynentalnej Azji Południowej. Przedstawiamy tutaj, o ile nam wiadomo, pierwszy jednoznaczny okaz Homo ze środkowego plejstocenu z kontynentalnej Azji Południowo-Wschodniej i omawiamy jego taksonomiczne przypisanie i implikacje dla ewolucji człowieka w tym regionie.W grudniu 2018 roku, stały dolny ząb trzonowy hominina został odnaleziony z bloku brekcji w Tam Ngu Hao 2 (Jaskinia Cobra), prowincja Huà Pan, Laos (20°12′41.5′′N, 103°24′32.2′′E, wysokość 1116 m; Rys. 1 , Rys. uzupełniający 1 ). Kras wieżowy, w którym uformowała się jaskinia, znajduje się po południowo-wschodniej stronie góry P’ou Loi, a wejście znajduje się 34 m nad równiną aluwialną (Rys. 1a , Rys. uzupełniający 1 ). Miejsce to zostało odkryte podczas badania obszaru wokół Tam Pà Ling, gdzie wcześniej odnaleziono skamieniałości wczesnego Homo sapiens 20 , 21 , 22 . Ząb (TNH2-1) jest zawiązkiem korony lewego stałego zęba trzonowego żuchwy (Ryc. 2a–f ; Ryc. uzupełniająca 2 ), a brak zużycia okluzyjnego i międzyzębowego w połączeniu z początkowym formowaniem się korzenia sugeruje, że ząb nie wyrósł w momencie śmierci osobnika. Morfologia zęba jest zgodna z przypisaniem do pierwszego lub drugiego dolnego zęba trzonowego (Metody – szczegółowa analiza morfologiczna zęba). W obu przypadkach, biorąc pod uwagę wczesny etap dojrzewania korzenia, ząb ten należał do młodego osobnika odpowiadającego wiekowi od 3,5 do 8,5 roku zgodnie z nowoczesnymi standardami rozwojowymi 23 .

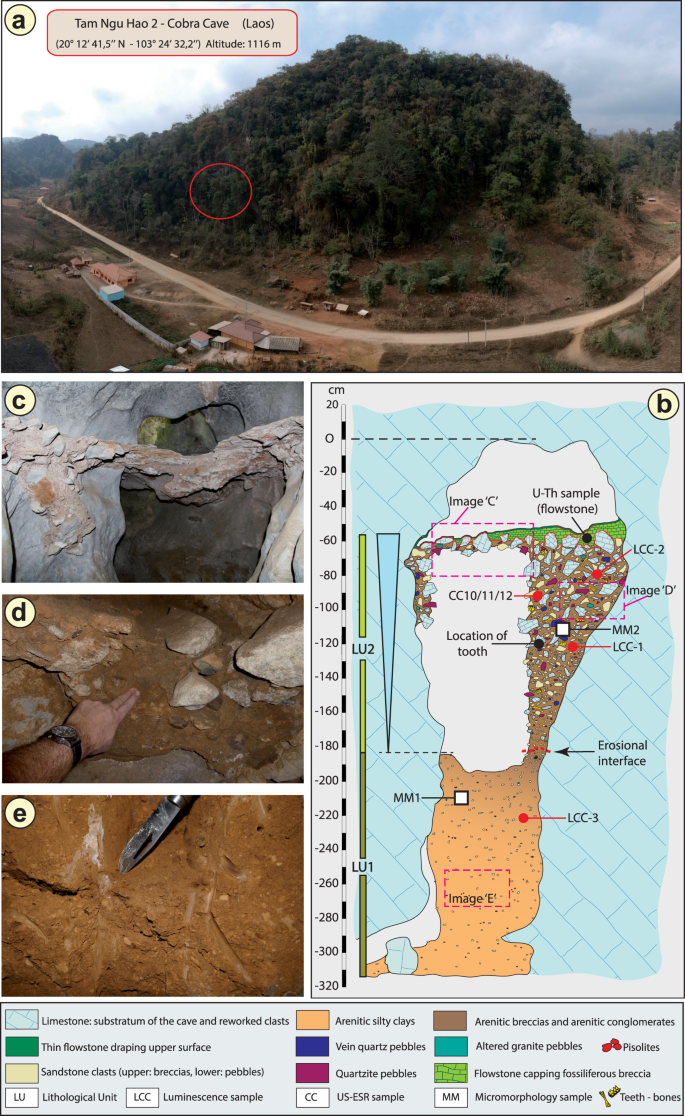
a Widok z lotu ptaka na stanowisko. Czerwone kółko wskazuje wejście do jaskini Tam Ngu Hao 2. b Stratygrafia i miejsca poboru próbek wypełnienia jaskini, pokazujące jednostki litologiczne 1 i 2 (LU1 i LU2) z erozyjnym interfejsem między tymi warstwami oznaczonym przerywaną czerwoną linią; Pokazano również próbki mikromorfologiczne (mikrostratygraficzne) (MM1 i MM2). Zakreślone liczby oznaczają przybliżone pozycje fotografii w c , d i e . c Widok nacieku wieńczącego górną pozostałą część LU2. d Szczegół brekcji/zlepieńca arenitowego LU2. e Szczegół iłu arenitowego LU1.

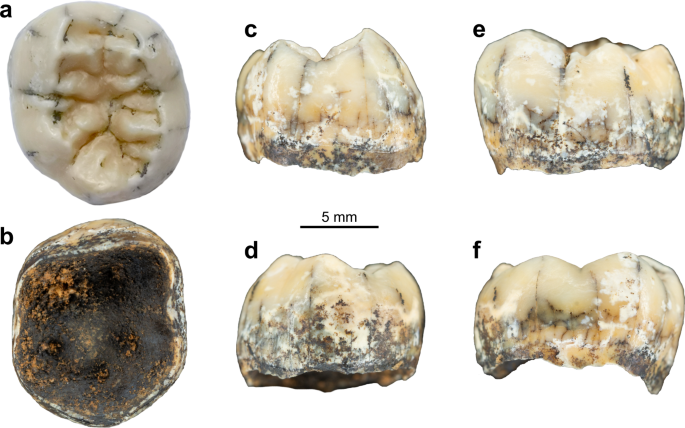
Zdjęcia TNH2-1 w widoku zwarciowym ( a ), dolnym ( b ), mezjalnym ( c ), dystalnym ( d ), policzkowym ( e ) i językowym ( f ).
W tej pracy, aby jak najlepiej udokumentować THN2-1, wykonano opis morfologiczny i analizy porównawcze. Opracowaliśmy również specjalny protokół pobierania próbek, który pozwala nam pobierać próbki do analiz paleoproteomicznych i przyszłych analiz izotopowych, zachowując jednocześnie całą morfologię powierzchni okluzyjnej korony. Pobieranie próbek do tych analiz destrukcyjnych odbywa się po analizie mikroCT całego zęba, zapewniając zachowanie pełnych danych morfologicznych. Na tym etapie nie wykonuje się żadnego dodatkowego pobierania próbek do analiz starożytnego DNA, biorąc pod uwagę wiek okazu i tropikalne warunki, w których osad i skamieliny zostały zdeponowane. Inwazyjna strategia pobierania próbek w celu zebrania tkanek zębowych do analiz molekularnych koncentruje się tylko na dystalnej części dolnej części korony, zachowując nienaruszoną część mezjalną korony.
Wyniki
Kontekst i datowanie
Przeanalizowano środowisko geologiczne, stratygrafię i mikromorfologię sekwencji osadów, aby uzyskać kompleksową, wieloskalarną ocenę kontekstu depozycyjnego i historii tafonomicznej skamieniałości wydobytych z jaskini (Informacje uzupełniające, Geologia). Częściowo zerodowane osady wypełniające badany korytarz wejściowy składają się z dolnej i górnej facji reprezentujących dwie fazy akumulacji osadów oddzielone powierzchnią erozyjną i nieznanym okresem czasu (Rys. 1b ). Dolna facja (jednostka litologiczna 1, LU1) jest słabo scementowana i tworzy arenitowe osady ilaste pozbawione skamieniałości (Rys. 1e ). Górna, skamieniała facja (jednostka litologiczna 2, LU2) jest dobrze scementowana i gruboziarnista, zawiera wewnątrzkrasowe kanciaste klasty wapienia i pozakrasowe zaokrąglone otoczaki, tworząc bardzo twardą warstwę brekcji/zlepieńca, z której elementy szkieletowe — a w szczególności zęby — zostały odzyskane w wysokiej częstotliwości (rys. 1d ). Zmiana litologii między dwiema facjami najprawdopodobniej odzwierciedla rekonfigurację krasowego systemu hydrologicznego, co byłoby związane z dużą powodzią, erodującą przestrzeń w LU1, na którą osady LU2 zostały niezgodnie nałożone. Osady LU2 są bocznie ciągłe i gęsto upakowane w całym odsłonięciu wykopanym na potrzeby tego badania, co uniemożliwia większą przeróbkę materiału i potwierdza kontekst stratygraficzny skamieniałości zawartych w nim, w tym zęba hominina (patrz szczegółowe obserwacje opisane w Metodach). Górna facja (LU2) jest usiana dwoma węglanowymi naciekami, co wskazuje na ostateczną zmianę hydrologii i odpływ wód powierzchniowych z jaskini oraz wytrącanie się laminarnego speleothemu (ryc. 1c ).
Trzy zęby wołowe (TNH2-10/CC10, TNH2-11/CC11, TNH2-12/CC12) wydobyte z górnej brekcji zawierającej skamieniałości (LU2) datowano bezpośrednio za pomocą sprzężonych szeregów uranowych i rezonansu spinowego elektronów (US-ESR), uzyskując ważoną średnią ocenę wieku na poziomie 151 ± 37 tysięcy lat temu (tys. lat) (2-sigma) (rys. 1b ; tabele uzupełniające 1 , 2 ) i przedział wieku 188–117 tys. lat. Dwa duże bloki brekcji (LCC1 i LCC2) z LU2 (góra) i jeden blok jednostki iłu mulistego (LCC3) z LU1 (dół) pobrano w celu datowania luminescencyjnego (rys. 1b ). Próbki te dały równowiekowe szacunki wieku 143 ± 24 tys. lat (LCC1) i 133 ± 19 tys. lat (LCC2) dla osadzania brekcji LU2 i 248 ± 31 tys. lat (LCC3) dla leżącego niżej osadu iłu mulistego LU1 (Tabela uzupełniająca 3 ). Wiek ten jest zgodny ze stratygraficznym wiekiem wyżej położonego nacieku (CCF1), który wytrącił się wcześniej niż 104 ± 27 tys. lat na podstawie ważonej średniej szacunków wieku serii U dla czterech oddzielnych podpróbek węglanu nacieku (Tabela uzupełniająca 4 ). Modelowanie bayesowskie zostało przeprowadzone dla wszystkich niezależnych szacunków wieku w celu określenia ogólnych ram geochronologicznych dla stanowiska i zęba (Metody i Ryc. uzupełniająca 3 ). Brekcja zawierająca skamieniałości, w tym ząb, została zdeponowana w wieku od 164 do 131 tys. lat (przy granicy ufności 68%).
Fauna
Zespół faunistyczny Tam Ngu Hao 2 obejmuje 186 zidentyfikowanych okazów dentognatycznych (NISP), w których dominują izolowane zęby dużych ssaków, w tym kilku megaroślinożerców (Tabela uzupełniająca 5 , Dane uzupełniające 2 ). Ich analizy ujawniają typowe ścieżki tafonomiczne zespołów z systemów krasowych pod względem reprezentacji okazów i rodzajów uszkodzeń. Ze względu na energię związaną z depozycją LU2, w zespole obecne są tylko zęby dużych ssaków, a my zauważamy brak małych i lekkich zębów jakichkolwiek mikrokręgowców. Co więcej, większość zębów jest obgryzana przez jeżozwierze, znane jako główny czynnik akumulujący w tym regionie 24 . Dlatego też słabe zachowanie okazów uniemożliwia identyfikację do poziomu gatunku dla większości zarejestrowanych taksonów. Fauna wykazuje bliskie pokrewieństwo z tą znaną z późnego środkowego plejstocenu południowych Chin i północnych Indochin, a w mniejszym stopniu z Jawy, co jest zgodne z chronologią sedymentacyjną tego stanowiska. Można ją przypisać do „ kompleksu faunistycznego Stegodon-Ailuropoda ” 25 , 26 , 27 , 28 . Zauważamy brak taksonów neogenu, które przetrwały we wczesnym plejstocenie, oraz brak dwóch kluczowych gatunków, Pachycrocuta brevirostris i Gigantopithecus blacki , które są dobrymi wskaźnikami faun sprzed 300 tys. lat w tym regionie 26 , 27 , 29 , 30 , 31 . Archaiczny Stegodon przetrwał w Azji najprawdopodobniej do końca późnego plejstocenu 32 . Odkryliśmy roślinożerców, w tym Tapirus , Stegodon i Rhinocerotidae, które przystosowały się do zadaszonych lasów w tym rejonie. Znaleźliśmy również zwierzęta, takie jak gatunki Bos , małe Caprinae i duże Cervidae (prawdopodobnie Rusa unicolor ), które są znane z dużej zmienności w zachowaniu żywieniowym i preferowanych siedliskach, od zamkniętych i przejściowych lasów po otwarte łąki33 .
Analizy starożytnych białek
Szkliwo z próbki zęba TNH2-1 zostało przeanalizowane przy użyciu nanoLC-MS/MS i niedawno opracowanego podejścia do starożytnych proteomów szkliwa 34 . Proteom TNH2-1 składa się ze wspólnego zestawu specyficznych dla szkliwa białek, z których wszystkie były wcześniej obserwowane w plejstoceńskich proteomach szkliwa 34 , 35 , 36 (Tabela uzupełniająca 6 ). Proteom szkliwa ma podwyższone poziomy diagenetycznych modyfikacji białkowych (Rysunek uzupełniający 4a–d , Tabela uzupełniająca 7 ) i zachowuje fosforylację seryny (S) w motywie SxE wcześniej obserwowanym w starożytnym szkliwie zębów 34 , 35 (Rysunek uzupełniający 4e ). Biorąc pod uwagę skład i modyfikację proteomu, a także brak peptydów pasujących do któregokolwiek z tych białek w naszych próbkach ekstrakcyjnych i spektrometrii masowej, uważamy, że nasze dane proteomiczne wskazują na obecność białek endogennych pochodzących z pobranych próbek szkliwa.
Niestety, żadne peptydy o wysokim poziomie pewności nie nakładały się na diagnostyczne pozycje aminokwasów z różnicami sekwencji między H. sapiens , Denisowianami lub Neandertalczykami, co uniemożliwiało dalsze przypisanie taksonomiczne oparte na paleoproteomice. Jest to zgodne z wcześniejszymi badaniami, które wskazywały, że blisko spokrewnione populacje homininów można odróżnić na podstawie proteomów zębiny i kości, podczas gdy proteomy szkliwa są mniej informacyjne w kontekście bliskiej bliskości filogenetycznej 35 . Niemniej jednak, porównując sekwencje odzyskane z proteomu szkliwa TNH2-1 z sekwencjami współczesnych hominidów, dla których dostępne są sekwencje białkowe, stwierdzamy, że okaz należy do członka rodzaju Homo (Tabela uzupełniająca 8 ).
Brak peptydów specyficznych dla męskiej amelogeniny Y (AMELY) sugeruje, że albo pobrany próbka zęba trzonowego pochodziła od osobnika płci żeńskiej, albo że nie zaobserwowano peptydów specyficznych dla AMELY z powodu degradacji wykraczającej poza granicę wykrywalności urządzenia.
Analizy strukturalne zewnętrzne i wewnętrzne zęba
Zewnętrznie korona TNH2-1 wykazuje gruby wzór zmarszczek, który występuje u Homo plejstoceńskiego ( H. erectus sl, europejscy i azjatycki Homo środkowoplejstoceńscy oraz neandertalczycy), ale jest rzadki u współczesnego H. sapiens . Środkowy grzebień trygonidowy jest dobrze rozwinięty, jak powszechnie odnotowano u europejskiego Homo środkowoplejstoceńskiego i neandertalczyków, podczas gdy jest on ogólnie nieobecny lub występuje rzadziej u H. erectus sl oraz kopalnych i współczesnych H. sapiens 37 . Poniżej zewnętrznej powierzchni, połączenie szkliwa z zębiną (EDJ) zęba pokazuje rogi zębiny pięciu głównych guzków i guzka pośredniego oraz niski, ale nieprzerwany środkowy grzebień trygonidowy (ryc. 2 i 3 , Ryc. uzupełniający 2 ; Metody). Ta ostatnia cecha jest ogólnie spotykana u neandertalczyków (80–100% w zależności od położenia zęba trzonowego) 38 , 39 , 40 , ale jest rzadsza u H. erectus sl i H. sapiens41 , 42 , 43 , 44 , 45 , 46 , 47 (Rys. uzupełniający 5 ) . Ponadto, EDJ TNH2-1 pokazuje wewnętrznie umieszczony metakonid przypominający zęby trzonowe neandertalczyków40 i niską topografię korony podobną do tej u H. erectus41 , 42 , 43 , 44 , 45 , 46 , 47 . Cechy te, jak również niewielka półka policzkowa obecna na EDJ TNH2-1, są wyrażone na EDJ zębów trzonowych denisovańskich z jaskini krasowej Baishiya (Xiahe, Gansu, Chiny) (Rys. uzupełniający 5 ) 15 . Zębina TNH2-1 różni się od znacznie wyższej i proporcjonalnie bardziej mezjodystalnie ściśniętej EDJ neandertalczyków i H. sapiens 39 , 40 , a także od krótszych rogów zębiny i gęściej pomarszczonej niecki zwarciowej H. erectus sl 41 , 42 , 43 , 44 , 45 , 46 , 47 (Rys. uzupełniający 5 ).

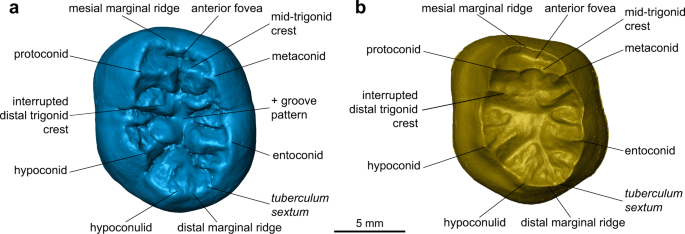
Wirtualne wizualizacje zewnętrznej powierzchni szkliwa ( a ) i połączenia szkliwa z zębiną ( b ) w widoku okluzyjnym, pokazujące główne cechy morfologiczne.
Jeśli chodzi o wymiary bezwzględne, tylko azjatycki Homo ze środkowego plejstocenu ma większe korony zębów niż TNH2-1 (tabele uzupełniające 9 , 10 ). Parametry koron TNH2-1 mieszczą się w zakresach zmienności dla H. erectus sl, H. antecessor , azjatyckiego Homo ze środkowego plejstocenu i neandertalczyków, ale statystycznie różnią się od mniejszych koron europejskiego Homo ze środkowego plejstocenu oraz od plejstocenu i holocenu H. sapiens (ryc. 4 ; tabele uzupełniające 10 , 11 ). Jeśli chodzi o proporcje tkanki korony zęba, TNH2-1 ma wysoki odsetek zębiny koronowej (Vcdp/Vc: 55,37%) ze średniogrubym szkliwem, jak pokazują wartości bezwzględnej i względnej grubości szkliwa (3D AET: 1,18 mm; 3D RET: 17,00; tabela uzupełniająca 12 ). Te proporcje tkanki korony odpowiadają proporcjom prawie niezniszczonego M2 żuchwy Xiahe 15 (Vcdp/Vc: 54,62%; 3D AET: 1,47 mm; 3D RET: 18,97) i górnego zęba trzonowego Denisova 4 (3D RET: 15,27; B. Viola, pers. comm.), ale mieszczą się w zakresach zmienności wszystkich porównywalnych kopalnych i współczesnych grup ludzkich (Rys. uzupełniający 6a–c ; Tabele uzupełniające 12 , 13 ). Trójwymiarowe mapy topograficznego rozkładu grubości szkliwa pokazują, że TNH2-1 ma najgrubsze szkliwo na szczycie guzków hipokonidalnych i hipokonulicznych oraz w dystalnej ćwiartce policzkowej korony (Rys. uzupełniający 6d ). Dla porównania, wszystkie inne próbki mają tendencję do posiadania najgrubszej emalii rozłożonej na wszystkich guzkach policzkowych i bardziej rozłożonej na policzkowej stronie korony, nawet jeśli jest ona zmienna między grupami i między pozycjami zębów trzonowych. M2 próbki Xiahe pokazuje grubszą emalię rozłożoną wzdłuż policzkowej strony korony, ale jej wzór rozłożenia jest częściowo zatarty przez zużycie okluzyjne.


Dwuwymiarowe wykresy rozrzutu wymiarów korony mezjodystalnej i policzkowo-językowej TNH2-1 w porównaniu z M1 ( a ) i M2 ( b ) H. erectus (HE; zielone krzyżyki), H. antecessor (HA; zielone plusy), środkowoplejstoceńskich neandertalczyków (MNEA; pomarańczowe trójkąty), późnoplejstoceńskich neandertalczyków (LNEA; czerwone odwrócone trójkąty), azjatyckich środkowoplejstoceńskich Homo (AMPH; ciemne kropki), późnoplejstoceńskich współczesnych ludzi (LPMH; niebieskie gwiazdki) i holoceńskich ludzi (HH; niebieskie romby). Dane źródłowe są dostarczane jako plik Source Data.
Kształt EDJ TNH2-1 porównano ilościowo z kształtami plejstoceńskich i holoceńskich grup ludzkich przy użyciu geometrycznej morfometrii (metody). Zastosowano podejścia oparte na punktach orientacyjnych i deformacjach powierzchni, przy czym obie metody w podobny sposób rozróżniały H. erectus sl, europejskich środkowoplejstoceńskich Homo i neandertalczyków oraz H. sapiens przy użyciu analizy zmiennych kanonicznych i głównych składowych między grupami (ryc. 5 , ryc. uzupełniająca 7 ). Wzdłuż CV2 i bgPC1, wyższe EDJ i bardziej zewnętrznie osadzone rogi zębiny neandertalczyków i H. sapiens są odróżniane od niższych i bardziej centralnie położonych rogów zębiny zębów trzonowych H. erectus . Osie CV1 i bgPC2 oddzielają neandertalczyków od współczesnych ludzi, przy czym pierwsi mają więcej wewnętrznie umieszczonych rogów zębiny mezjalnej i bardziej rozwinięty hypoconulid niż drudzy. TNH2-1 nie mieści się w zakresach wszystkich innych grup. Ma kształt pośredni EDJ pomiędzy niską koroną H. erectus (ale przewyższający zmienność tej ostatniej grupy wzdłuż CV1 i bgPC2) a pozycją guzka zębów trzonowych neandertalczyków (nawet jeśli znajduje się poza zakresem ich zmienności wzdłuż CV2 i bgPC1). Najbliższe powinowactwo morfologiczne TNH2-1 leży w okazie denisowiańskim Xiahe, który również wykazuje cechy neandertalskie (ryc. 5 , ryc. uzupełniająca 7 ).

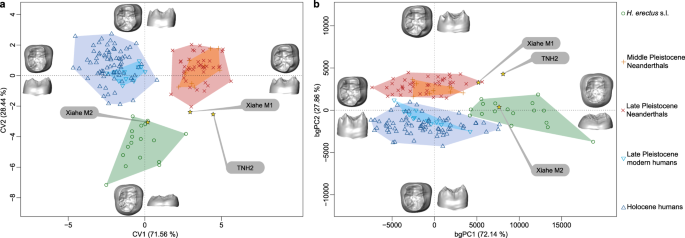
Analiza zmiennych kanonicznych ( a ) i analiza głównych składowych między grupami ( b ) porównania kształtu TNH2-1, H. erectus sl, okazu denisowiańskiego z Xiahe, neandertalczyków i H. sapiens na podstawie deformacji EDJ . Dane źródłowe podano w postaci pliku Source Data .
Dyskusja
Rekonstrukcja rozproszenia i ostatecznie trajektorii ewolucyjnych Homo w Azji zależy od obecnie ubogiego zapisu kopalnego. Azjatycki zapis kopalny z późnego środkowoplejstocenu jest w większości ograniczony do wschodniej części kontynentu 4 , 8 , 9 , 10 , 15 , 48 . Wszelkie dodatkowe szczątki ludzkie z tego okresu dokumentujące ewolucję Homo w południowej Azji mogą zatem pomóc potwierdzić wcześniejsze hipotezy lub ujawnić nowe linie rozwojowe. Analiza proteomiczna zęba trzonowego TNH2-1 wskazuje, że najprawdopodobniej należy on do osobnika żeńskiego z rodzaju Homo . Analizy morfometryczna zewnętrznej i wewnętrznej organizacji strukturalnej korony pozwalają nam odrzucić szereg hipotez dotyczących przypisania gatunku. TNH2-1 ma duże wymiary korony i złożoną powierzchnię zgryzową, co odróżnia go od mniejszych i morfologicznie prostszych zębów H. floresiensis 49 , H. luzonensis 50 i H. sapiens (ryc. 4 ). Kształt EDJ wykazuje mieszankę cech neandertalskich i H. erectus , ściśle przypominając morfologię M1 okazu denisowiańskiego z Xiahe (Ryc. 5 , Ryc. uzupełniający 5 ). Podobieństwa między TNH2-1 i H. erectus są głównie związane z proporcjonalnie niższą koroną, chociaż trzonowce H. erectus wykazują jeszcze niższe korony trzonowców i węższą nieckę zwarciową (Ryc. 2 i 3 , Ryc. uzupełniający 5 ). Skamieniałość laotańska wykazuje wyraźne cechy neandertalskie, takie jak dobrze rozwinięty środkowy grzebień trygoniczny i wewnętrznie ułożone mezjalne rogi zębiny, ale różni się znacznie niższą topografią EDJ i kształtem niecki zwarciowej.
Różnice w stosunku do neandertalczyków, które obserwujemy, nie wykluczają przynależności TNH2-1 do tego taksonu i uczyniłyby go najbardziej wysuniętą na południowy wschód skamieniałością neandertalczyka, jaką kiedykolwiek odkryto. Jednakże, biorąc pod uwagę morfologiczne osobliwości TNH2-1 w unisonie, a także wysoki stopień podobieństwa morfologiczno-wymiarowego do zębów trzonowych okazu denisowiańskiego z Xiahe, najbardziej oszczędną hipotezą jest to, że TNH2-1 należy do tej siostrzanej grupy neandertalczyków. Jeśli TNH2-1 rzeczywiście należy do denisowianina, to wystąpienie, wraz z niedawnym odkryciem żuchwy denisowianina z Wyżyny Tybetańskiej, wysokogórskiego, niedotlenionego środowiska 15 , sugerowałoby, że ta plejstoceńska populacja azjatycka posiadała wysoki stopień plastyczności, aby przystosować się do bardzo zróżnicowanych środowisk 51 . Dostępne szczątki zębowe denisowian wskazują na mieszankę cech zgodnych z obecnymi dowodami paleogenetycznymi, że denisowianie i neandertalczycy są taksonami siostrzanymi 13 , 14 , 51 , 52 , 53 i dlatego oczekuje się, że będą mieć pewne wspólne cechy czaszkowo-zębowe 15 , 54 . Potwierdzają to również niedawne analizy, które zidentyfikowały możliwe cechy szkieletu denisowian na podstawie jednokierunkowych zmian metylacji, w tym cechy, które zostały powiązane z chińskimi skamieniałościami, takimi jak Xujiayao i Xuchang 9 , 54 . Denisowianie wyróżniają się dużym uzębieniem z pewnymi cechami korony przypominającymi neandertalczyka 15 , 48 , 54 , a także charakterystyczną morfologią guzków i korzeni 14 , 15 , 16 . W obliczu braku analiz molekularnych poszukiwanie tych połączonych cech w azjatyckich skamieniałościach człowieka, w tym w skamieniałościach takich jak żuchwa Penghu 1 z Cieśniny Tajwańskiej55 , może pomóc w identyfikacji większej liczby okazów denisowiańskich (Rys. uzupełniający 8 ).
Hipotezy alternatywnej, zgodnie z którą TNH2-1 należy do grupy neandertalczyków, którzy dokonali inwazji na Azję Południowo-Wschodnią (zob. na przykład dyskusje na temat skamieniałości, które mogą świadczyć o tym rozproszeniu z Maba i Dali) 56 , 57 nie można całkowicie odrzucić, nawet jeśli jest ona mniej prawdopodobna.
Ząb z jaskini Tam Ngu Hao 2 w Laosie dostarcza zatem bezpośredniego dowodu na istnienie najprawdopodobniej samicy denisowiańskiej z towarzyszącą jej fauną na kontynencie Azji Południowo-Wschodniej w wieku 164-131 tys. lat. Odkrycie to dodatkowo potwierdza, że region ten był punktem zapalnym różnorodności dla rodzaju Homo (Rys. uzupełniająca 8 ), z obecnością co najmniej pięciu gatunków z późnego środkowego do późnego plejstocenu: H. erectus 58, denisowianie/neandertalczycy, H. floresiensis 49 , H. luzonensis 50 i H. sapiens 20 , 21 , 22 .
Metody
Brekcję zawierającą skamieniałości wydobyto mechanicznie ze ścian jaskini, a niektóre z najtwardszych bloków zawierających faunę rozpuszczono w 10% roztworze kwasu mrówkowego.
Wykopaliska, zbiórka i eksport skamieniałości do badań zostały przyznane przez Ministerstwo Informacji, Kultury i Turystyki Laotańskiej Republiki Ludowo-Demokratycznej w styczniu 2019 r. przy wsparciu lokalnych władz dystryktu Xon, prowincji Hua Pan i mieszkańców wioski Long Gua Pa. Wyniki tego badania zostaną udostępnione władzom dystryktu Xon i mieszkańcom wioski Long Gua Pa zgodnie z tą samą procedurą informacyjną, którą ustaliliśmy na początku każdej z naszych corocznych prac terenowych.
Analiza mikrostratygraficzna
W celu analizy mikrostratygrafii osadów Cobra Cave, nienaruszony blok osadu został usunięty z LU1 (MM1) i LU2 (MM2) (rys. 1b ). Nieskonsolidowaną próbkę (MM1, LU1) wyekstrahowano za pomocą bandaża nasączonego gipsem w celu ustabilizowania osadu. Możliwe było usunięcie próbki z LU2 (MM2) za pomocą młotka i dłuta ze względu na dobrze scementowaną naturę brekcji. Próbka z LU2 została zlokalizowana w obszarze blisko (~30 cm) miejsca znalezienia zęba hominina. Cienkie sekcje przygotowano w Adelaide Petrographics (Adelaide, Australia). Blok z LU2 nie wymagał stabilizacji i został pocięty za pomocą piły do skał. Plaster o grubości ~100 µm został zamontowany na szkiełku przedmiotowym i oszlifowany i wypolerowany do końcowej grubości 30 µm. Blok osadu z dolnej nieskonsolidowanej jednostki zaimpregnowano żywicą, postępując zgodnie z procedurami opisanymi gdzie indziej 59 , 60. Cechy diagnostyczne zaobserwowane za pomocą mikroskopu polaryzacyjnego rejestrowano dla każdej warstwy, stosując standardowe protokoły 61 .
Mikromorfologiczne (mikrostratygraficzne) analizy wypełnień osadowych jaskiń mogą wyjaśnić procesy odpowiedzialne za formowanie się stanowisk skamieniałości, środowiska depozycyjne i postdepozycyjne oraz kontekst zespołów skamieniałości 59 , 60 , 62 , 63 , 64. Mikrokontekstualizacja może dostarczyć kluczowych informacji dotyczących kontekstu skamieniałości i tego, jak daleko mogły się one przemieścić od swoich pierwotnych pozycji.
LU1: Niższe piaski i muły (rys. uzupełniający 9a )
Niższe osady LU1 wydawały się piaszczyste i ziarniste w terenie, ale analizy mikrostratygraficzne pokazują, że inkluzje wielkości piasku składają się z ziaren kruszywa ilastego ilastego o zmiennym składzie litologicznym. Te niższe osady są na ogół luźne, ale z częściowym cementowaniem w zlokalizowanych strefach. Ogólnie rzecz biorąc, osady LU1 (Rys. uzupełniająca 9a ) składają się z i) materiału matrycowego z drobnego piasku kwarcowego w plamistej tkaninie B oraz ii) dużych, grubych, wielkości piasku, zaokrąglonych do dobrze zaokrąglonych ziaren kruszywa o zmiennym kolorze, litologii i układzie tkaniny, co sugeruje poligenetyczne (wewnątrz- i pozakrasowe) pochodzenie.
Groundmass waha się od pojedynczego odstępu do podwójnego odstępu enaulicznego. W skali makro, ziarna złożone tworzą grubą frakcję całego osadu, wahającą się od bardzo drobnego żwiru/bardzo grubego do drobnego piasku i w kształcie od kanciastego do dobrze zaokrąglonego. Gruby składnik materiału matrycy obejmuje bardzo drobny piasek kwarcowy i gruby muł, podkanciasty do podzaokrąglonego. Wiele z tych ziaren agregatu wykazuje wewnętrzne rozwarstwienie z mikropasmami drobniejszego i grubszego materiału (głównie gliny i mułu), a ziarna różnią się kolorem od jasnobrązowego do bardzo ciemnobrązowego. Osady są umiarkowanie porowate, wykazując częste płaskie, vughy i pęcherzykowe puste przestrzenie. Płaskie puste przestrzenie zazwyczaj łączą się pionowo lub podpionowo przez vughy i pęcherzyki i są powszechne w sąsiedztwie krawędzi dużych ziaren złożonych. Uwarstwione ziarna złożone są zazwyczaj znacznie mniej porowate i często są wzbogacone w glinę. Pedofeatures są rzadkie w LU1 i najczęściej widoczne są w postaci glinianych powłok wiszących otaczających zaokrąglone lub podzaokrąglone ziarna złożone. Małe cząstki żelaza występują w małych ilościach. Materiał organiczny jest nieobecny w tej warstwie.
Obserwacje terenowe i analizy mikromorfologiczne łącznie sugerują, że LU1 został zdeponowany w środowisku hydrologicznym o niskiej do umiarkowanej energii. Gruba natura ziaren kruszywa (bardzo drobny żwir, bardzo gruby piasek i sięgający do drobnego piasku) sugeruje, że woda przepływająca przez system krasowy przerobiła starsze wypełnienia jaskiń przylegające do ścian i stropu wyżej w systemie zlewni jaskini (por. 62 ). Zmienne litologie tych złożonych ziaren są zgodne z przeróbką drobnoziarnistego materiału z wielu źródeł, najprawdopodobniej pochodzenia zarówno wewnątrzkrasowego, jak i pozakrasowego. Bardzo ciemne zabarwienie niektórych (≤5%) ziaren kruszywa sugeruje albo przerobiony materiał pedogeniczny, albo pelit pochodzący spoza stanowiska. Po osadzeniu tych sterylnych skamieniałościowo osadów, wystąpiły niewielkie zaburzenia osadów po depozycji, o czym świadczy dobrze rozwinięta struktura pustki zgodna z bioturbacją. Bioturbacja przyczyniła się do usunięcia wszelkich pasm, warstw i struktur świadczących o specyficznych procesach sedymentacji rzecznej.
LU2: Górny konglomerat/brekcja zasobna w skamieliny (Rys. uzupełniający 9b–t )
Jak wspomniano powyżej, konglomerat pobrano w celu uniknięcia klastów żwirowych, więc ten opis dotyczy drobnej matrycy osadowej. Obecne są losowo rozmieszczone puste przestrzenie, z powszechnymi bardzo małymi wyrwami i płaskimi pustkami, które są lokalnie większe i bardziej złożone, czasami tworząc komory i pustki kanałowe. Wszystkie puste przestrzenie są wypełnione węglanem wapnia w celu scementowania osadu. Masa podstawowa jest na ogół podwójnie rozmieszczonym porfirowym wzorem rozkładu c/f z częstymi kątowymi do podkątowych ziarnami mineralnymi (głównie kwarcu), małymi kątowymi fragmentami kości, dużymi fragmentami zębów i kilkoma małymi (drobnymi do średnich wielkości piasku) grudkami zabarwionymi żelazem. Tkanina jest rzadko granostriowana wokół grubych inkluzji (np. ziaren kwarcu lub skał). Cechy pedo są rzadkie w osadach jaskini TNH2, ale rzadkie powłoki iłu są rejestrowane w niektórych obszarach badanego cienkiego przekroju, z cienkimi powłokami na niektórych fragmentach kości. Istnieją również obszary, w których matryca osadowa jest wzbogacona w glinę, prawdopodobnie w wyniku iluwiacji. Fragmenty kości są zazwyczaj średniej lub grubej wielkości piasku w cienkim przekroju (z większymi elementami odzyskanymi z osadu) i wykazują jedynie umiarkowane oznaki wietrzenia chemicznego, z niską dwójłomnością (w xpl) i pewnymi oznakami przebarwień tlenkowych prawdopodobnie związanych z aktywnością mikrobiologiczną. W niektórych obszarach próbki małe fragmenty kości są szczególnie wietrzejące i wykazują małe pęknięcia i silne utlenienie, a zatem mogą one reprezentować starsze, opóźnione osady. Obecne są duże fragmenty zębów (~15 mm), w tym zarówno zębina, jak i cement w dobrze zachowanym stanie.
Osady LU2 składają się z drobnych do średnich żwirów, gruboziarnistych do dużych klastów żwirowych i okazjonalnych otoczaków w kierunku górnego napływu. Żwir jest zawieszony w drobnoziarnistej matrycy mułów i drobnych do średnich piasków, ze zmienną ilością gliny. Osady te zostały osadzone w środowisku o wysokiej energii, prawdopodobnie podczas powodzi lub rekonfiguracji krasowego systemu hydrogeologicznego. Prawdopodobnie wydarzyło się to jako pojedyncze zdarzenie w ciągu kilku godzin lub dni, ponieważ nie identyfikujemy podjednostek, potencjalnie zarówno jako odpływ rzeczny, jak i nasycone przepływy gruzu. Ruchy masowe są rejestrowane w mikrostratygrafii jako granostriowane tkaniny b otaczające grube ziarna mineralne, dowód, że wokół ziaren zastosowano ciśnienie (rotacyjne). Osady te były prawdopodobnie nasycone wodą i plastyczne, a przesiąkająca woda przemieszczała drobne gliny przez profil, koncentrując te drobne w zlokalizowanych obszarach matrycy, jak odnotowano w zlokalizowanych obszarach badanych osadów.
Analizy mikromorfologiczne potwierdzają obserwacje terenowe, że małe fragmenty kości i częste duże fragmenty zębów są powszechne w całym badanym obszarze LU2, co wskazuje na długi czas przebywania w sieci krasowej. Fragmenty kości są zwykle kątowymi aspektami zatrzymującymi ich oryginalną morfologię i są ogólnie dobrze zachowane, z wyjątkiem umiarkowanego przebarwienia żelazem, co jest zgodne z działaniem mikroorganizmów. Jeśli chodzi o sygnaturę diagenetyczną i tafonomiczną zarejestrowaną w tych osadach, istnieje niewiele dowodów na trawienie kwasem lub zaokrąglanie krawędzi składników organicznych, chociaż niektóre ziarna kwarcu wykazują cechy wietrzenia. Tylko bardzo niewiele fragmentów kości (<5%) wykazuje oznaki znaczącego wietrzenia, co jest zgodne z dłuższym czasem przechowywania w sieci krasowej. Chociaż rejestrujemy bioturbację osadów na małą skalę, jest ona niewielka, a utworzone puste przestrzenie nie są wystarczająco duże, aby być źródłem mieszania osadów.
Randkowanie
Datowanie luminescencyjne matrycy brekcji
Dwa duże bloki twardej, cementowanej brekcji zawierającej skamieliny (LCC1 i LCC2) z LU2 i jeden z dolnej jednostki iłu mulistego (LCC3) w LU1 zostały wycięte z sekcji in situ i owinięte w czarny plastik (rys. 1b ). W warunkach przytłumionego światła czerwonego, zewnętrzna warstwa wystawiona na działanie światła została usunięta za pomocą dłuta i młotka i została zachowana jako próbka dozymetryczna. Warstwy te rozbito za pomocą tłuczka i moździerza i wysuszono w piecu, a następnie całą frakcję zmielono i wykorzystano do oszacowania dawki środowiskowej. Niewyeksponowany rdzeń wewnętrzny został również rozbity za pomocą tłuczka i moździerza i przetworzony przy użyciu standardowych procedur oczyszczania próbek w celu oddzielenia skalenia 65 , w tym 10% płukania w kwasie fluorowodorowym przez 10 minut w celu usunięcia zewnętrznych skórek alfa-dozowanych 58 . Wszystkie analizy luminescencji przeprowadzono w ośrodku datowania luminescencyjnego „Traps” na Uniwersytecie Macquarie w Sydney w Australii.
Pojedyncze ziarna skalenia o wielkości 180–212 µm umieszczono na powlekanych, jednoziarnistych dyskach w siatce 10 × 10. Dyski załadowano na karuzelę i poddano obróbce w urządzeniu Riso TL-DA-20 zawierającym zautomatyzowaną głowicę detekcyjno-stymulacyjną z podwójnym laserem jednoziarnistym i czułym na światło niebieskie/UV mikroskopem elektronowym PMT (PDM9107Q-AP-TTL-03) o maksymalnej wydajności detekcji w zakresie od 200 do 400 nm. Filtry w automatycznym zmieniaczu detekcji zostały ustawione na niebieskim pakiecie filtrów (filtry Schott BG-39 i Corning 7–59 do przesyłania długości fal 320–480 nm 66 . Ziarna były stymulowane za pomocą lasera modulowanego TTL IR (830 nm) 140 mW z filtrem długoprzepustowym RG-780 3 mm (zamontowanym bezpośrednio przed laserem IR), a emisje były wykrywane za pomocą kombinacji niebieskich filtrów opisanej powyżej. Laser stymulował ziarna przez 2,5 s, najpierw w 50 °C, a następnie w 270 °C, po wstępnym podgrzaniu do 300 °C zgodnie z procedurami protokołu pIR-IRSL 67 (patrz Testy proceduralne poniżej). Proces lokalizacji dysku pojedynczego ziarna został zaprogramowany tak, aby występował przed jakąkolwiek procedurą ogrzewania dysku, aby upewnić się, że każde ziarno otrzymało takie samo ogrzewanie w cyklach SAR.
Równoważne dawki skorygowano zgodnie z wynikami testów zanikania anomalii (stosując ważoną średnią szybkość zanikania wynoszącą 2,0 ± 0,2% na dekadę), a następnie przepuszczono przez model minimalnego wieku (MAM) w celu zidentyfikowania populacji, w której przed pochówkiem nastąpiło największe wybielenie.
Aby uzyskać oszacowanie mocy dawki środowiskowej dla każdej z próbek, najpierw zmierzyliśmy moce dawki beta, używając wielolicznika Geigera-Mullera do zliczania beta wysuszonych i sproszkowanych próbek osadów 68 w laboratorium. Uwzględniono wpływ zawartości wilgoci w próbce, różnych rozmiarów ziarna 69 i trawienia HF na tłumienie dawki beta, a całkowity wkład mocy dawki beta obliczono, porównując moc zliczania beta ze standardowym źródłem beta (SHAP o mocy dawki 5,99 Gy/kyr) i tlenkiem magnezu jako materiałem tła nieemitującym promieniowania beta. Po drugie, zliczanie alfa grubego źródła przy użyciu inteligentnego licznika alfa Daybreak 583 zostało użyte do uzyskania szacunków uranu i toru 70 w celu oszacowania mocy dawki gamma, a po trzecie, różnica między zliczaniem beta i alfa została użyta do oszacowania wartości potasu. Następnie te szacunki przeliczono na moce dawek gamma, używając współczynników konwersji Guerin i in. 71 . Uwzględniono wpływ wilgotności próbki 65 na zewnętrzne dawki promieniowania beta i gamma, stosując długoterminową zawartość wody wynoszącą od 3 ± 0,2 do 5 ± 2%, co jest podobne do zmierzonej (terenowej) zawartości wody wynoszącej od 3% do 7% i pozwala na początkowy okres nasycenia po pierwszym osadzeniu w środowisku krasowym.
Całkowitą moc dawki obliczono następnie, wykorzystując efektywną wewnętrzną moc dawki beta wynoszącą 0,84 Gy kyr −1 72 , 73 dla próbek skalenia o średnicy 180–212 µm (wskutek rozpadu promieniotwórczego 40 K i 87 Rb), które przygotowano, przyjmując stężenia K (12,5 ± 0,5%) i 87 Rb (400 ± 100 μg g –1 ) i uwzględniono w całkowitej mocy dawki. Moc dawek promieniowania kosmicznego oszacowano na podstawie opublikowanych zależności74 , biorąc pod uwagę grubość wapienia nad jaskinią (~20 m przy założonej gęstości 1,2 g/cm3 ) , nadkład osadów w miejscu poboru próbki (0,3–1,3 m przy założonej gęstości 2,0 g/cm3 ) , wysokość (~1116 m nad poziomem morza) oraz szerokość i długość geograficzną (20°S i 103°E) miejsca poboru próbki.
Ponadto, oszacowanie mocy dawki zostało poparte spektrometrią gamma o wysokiej rozdzielczości, aby zmierzyć aktywność radionuklidów w łańcuchach rozpadu 238 U, 235 U, 232 Th i 40 K w laboratorium SGS w Melbourne. Aktywności te zostały przeliczone na moce dawki beta i gamma przy użyciu współczynników konwersji Guerin et al. 71 i dostosowane do tej samej długoterminowej zawartości wody, o której wspomniano powyżej. Aktywności te zapewniają alternatywną dozymetrię i zostały również wykorzystane do oszacowania potencjalnej nierównowagi w tym środowisku jaskiniowym.
Testy proceduralne
Poprzednie analizy luminescencji w sąsiednim miejscu, Tam Pà Ling20 , 21 , 22 , ujawniły wysoką moc dawki (~3–4 Gy/kyr), nasycający sygnał kwarcowy o głębokości >4 m oraz skalenie o wysokiej odpowiedzi na dawkę i stabilnym sygnale, który można było odizolować od niestabilnego sygnału zanikającego22 . Opierając się na tym doświadczeniu, skupiliśmy się tylko na skaleniach i zastosowaliśmy ten sam protokół post-infra-red infra-red-stimulated-luminescence (pIR-IRSL), aby przezwyciężyć problemy anomalnego zanikania75 . Jednak ze względu na liczbę ziaren skalenia użytych na każdym dysku, co dało tylko maksymalne wieki osadzania osadów, w połączeniu z niską wydajnością ziaren skalenia potasowego w tych miejscach, zdecydowaliśmy się zastosować technikę pojedynczego ziarna pIR–IRSL zamiast techniki pojedynczego alikwotu.
Zastosowaliśmy te same testy proceduralne, które opisano w pracy Shackelford et al. 22 , ale zmodyfikowaliśmy je dla pojedynczych ziaren skalenia z próbki LCC1 (ponieważ ta próbka zawierała nieco więcej ziaren skalenia niż dwie pozostałe) zgodnie z protokołami Robertsa 76 i Rhodesa 77 (Tabela uzupełniająca 14 ): 1) test plateau wstępnego podgrzewania; 2) testy blaknięcia; 3) testy wybielania; i 4) testy odzysku dawki. Ze względu na niewielką ilość ziaren o wielkości frakcji 180–212 μm zachowaliśmy ją do oszacowania D e i zamiast tego użyliśmy mniejszej frakcji o wielkości 90–180 μm (i odpowiadających jej 200 μm pojedynczych krążków ziarnowych) do wszystkich testów proceduralnych, używając następujących kombinacji wstępnego podgrzewania i stymulacji IR: 1) 250 i 225 °C (pIR-IRSL 50,225 ) 67 , 78 ; 2) 280 i 250 °C (pIR-IRSL 50,250 ) 79 ; 3) 300 i 270 °C (pIR-IRSL 50,270 ) 79 ; 4) 320 i 290 °C (pIR-IRSL 50,290 ) 80 , 81 ; i ponieważ oczekiwane De jest >450 Gy, przetestowaliśmy również 5) 320 i 200/290 °C (pIR-IRSL 200.290 ) 82 . Ponieważ wiele dysków miało bardzo mało rozpadów skalenia, użyliśmy 4 dysków z pojedynczymi ziarnami o łącznej liczbie 16 ziaren i poddaliśmy te ziarna recyklingowi we wszystkich testach. Nie jest to rozwiązanie idealne i użycie świeżych ziaren byłoby lepsze, ale niska wydajność skalenia oznaczała, że musieliśmy nadać priorytet pozostałej próbce do rzeczywistych pomiarów. Wszystkie testy w większości przebiegały zgodnie z procedurami opisanymi w Shackelford et al., 22 z następującymi różnicami:
-
1.Testy plateau wstępnego nagrzewania i odzyskiwania dawki — różne kombinacje wstępnego nagrzewania/stymulacji laserem IR zastosowano do każdego dysku SG i do innej liczby ziaren, w zależności od tego, ile ziaren uległo luminescencji na każdym dysku. Następnie dodaliśmy dawkę zastępczą 20 Gy, aby spróbować odzyskać tę znaną dawkę z tych dysków.
-
2.Testy wybielania — te same 4 dyski poddano działaniu dawki 20 Gy i wybielono w symulatorze słonecznym, a następnie mierzono przy użyciu tych samych kombinacji wstępnego nagrzewania i stymulacji laserowej na podczerwień.
-
3.Postępując zgodnie z protokołami Rhodes 77 dotyczącymi testowania anomalnego zanikania w SG skaleni, zastosowaliśmy 1) indywidualne zanikanie pojedynczego ziarna przy użyciu 4 dysków przy wielokrotnym czasie opóźnienia; 2) wielokrotne pomiary ziarna przy użyciu dysków SG jako pojedynczych alikwot; i 3) zastosowanie standardowej oceny wartości zanikania na podstawie stopnia nasycenia dla nieskończenie starej próbki pochodzącej z zwietrzałych granitowych otoczaków znalezionych w złożu i analizowanych przy użyciu małych pojedynczych alikwot.
Testy wykazały, że najbardziej płaskie plateau zapewniały sygnały pIR-IRSL 50,270 , pIR-IRSL 50,290 i pIR-IRSL 200,290 (Rys. uzupełniający 10c ), podczas gdy sygnał pIR-IRSL 50,270 zapewniał najlepszą regenerację dawki zastępczej (ze współczynnikiem regeneracji dawki 0,995) i najniższe wartości resztkowe po wybielaniu (<10 Gy) (chociaż obliczone, te dawki resztkowe nie zostały odjęte od D e ) (Rys. uzupełniający 10e ). Sygnały pIR-IRSL 50,290 i pIR-IRSL 200,290 wytwarzały największe dawki resztkowe, co budzi obawy o przeszacowanie sygnału w tym środowisku jaskiniowym, w którym istnieje duże prawdopodobieństwo częściowego wybielania. Testy zanikania z testów pojedynczych alikwot były znacznie niższe niż z testów pojedynczych ziaren, które ujawniły zakres zanikania od najwyższego IR 50 do najniższego sygnału pIR-IRSL 50,270 , pIR-IRSL 50,290 i pIR-IRSL 200,290 (rys. uzupełniający 10d ). Zaobserwowaliśmy również zmienność szybkości zanikania wśród pojedynczych ziaren (od wartości ujemnych do wyższych wartości, które nie pozwoliłyby na żaden naturalny sygnał, ze średnią dla 16 ziaren wynoszącą 2,3% na dekadę), ale było to niezależne od czułości ziaren. Podczas używania sygnału pIR-IRSL 50,270 na dyskach pojedynczych alikwot, ta wartość g została zmniejszona do średniej 1,7% na dekadę. Postanowiliśmy użyć średniej szybkości zanikania wynoszącej 2,0 ± 0,2% na dekadę jako punktu środkowego przed dwiema technikami. Co zaskakujące, sygnał pIR-IRSL 200,290 wykazywał taki sam stopień zaniku, jak sygnały pIR-IRSL 50,270 i pIR-IRSL 50,290 , co wskazuje na brak wyraźnej przewagi wyższej początkowej temperatury przy stymulacji IR w tych próbkach.
Zwietrzały granitowy otoczak zapewnia najbliższą reprezentację pochodzenia ziaren skalenia z LCC1 i LCC2. Postępując zgodnie z procedurami Brown et al. 83 , użyliśmy 12 bardzo małych pojedynczych alikwot (ponieważ sygnał był zbyt nieczuły, aby użyć pojedynczych ziaren) i dodaliśmy duże dawki 200 i 400 Gy do naturalnego sygnału (N + 200 Gy i N + 400 Gy) dla ośmiu dysków i zerową dawkę dla ostatnich czterech dysków. Wszystkie dyski mierzono przy użyciu kombinacji pH/temperatury stymulacji opisanych powyżej (Rys. uzupełniający 10f ). We wszystkich dyskach sygnał IR 50 rośnie wraz z dawką, podobnie jak sygnał pIR-IRSL 50,225 w mniejszym stopniu, ale sygnały pIR-IRSL 50,290 i pIR-IRSL 200,290 zmieniają się tylko w niewielkim stopniu. Oznacza to, że nasycenie pola jest bliskie nasyceniu laboratoryjnemu przy użyciu tych sygnałów. Jednak, aby zachować ostrożność, zastosowaliśmy współczynnik zanikania wynoszący 2,0% na dekadę. Użyliśmy metody korekcji zanikania Huntleya i Lamothe84 , która zapewnia oszacowanie zanikania dla małych współczynników zanikania, chociaż jest zaprojektowana dla liniowej części krzywej wzrostu.
Ponieważ te skalenie zawierają bardzo duże wartości De blisko punktu nasycenia, przeprowadziliśmy testy nasycenia na tym samym zwietrzałym otoczaku, o którym wspomniano powyżej 83 . Najpierw uruchomiliśmy próbkę jako normalny pIR-IRSL 50,270 , aby zaobserwować odpowiedź dawki nasyconej próbki i upewnić się, że procedura dopasowania może uwzględnić te wysokie wartości dawkowania.
Komentarze do datowania luminescencyjnego brekcji
Skalenie miały niską wrażliwość dającą bardzo niskie wskaźniki akceptacji; łącznie przetworzono ponad 8300 ziaren, z których tylko 96 zostało zaakceptowanych (1,16% wskaźnik akceptacji). Frakcja wielkości 180–212 została wybrana do analiz pojedynczych ziaren, ponieważ była to największa dostępna frakcja wielkości; frakcja wielkości 90–125 dała nieistotne ziarna skalenia. Odrzucenia opierały się na protokole Jacobs et al. 85 (patrz tabela uzupełniająca 15 ). Ziarna zostały przetestowane za pomocą XRD w celu sprawdzenia, czy były to w rzeczywistości bogate w potas ziarna skalenia, a wyniki wskazały, że zawierały one nieco niższą niż oczekiwano zawartość potas (pomiędzy 10,9 a 12,5%). Większość ziaren w ogóle nie uległa rozkładowi. Te wyniki były trudne, biorąc pod uwagę, że niewielkie ilości próbek oznaczały, że nie byliśmy w stanie uzyskać liczby zaakceptowanych ziaren do statystycznie istotnej liczby przed wyczerpaniem próbki (LCC1 – 48 ziaren, LCC2 – 25 ziaren i LCC3 – 23 ziarna). Jednak wartość podejścia pojedynczego ziarna polega na tym, że możemy wyizolować poszczególne populacje dawek, a nie średnią wielu populacji, co skutkuje maksymalnym wiekiem. Tak więc oszacowanie wieku uzyskane metodą pojedynczego ziarna (nawet przy małej liczbie ziaren) jest znacznie bliższe rzeczywistemu wiekowi pochówku osadów niż podejście wielokrotnego ziarna, pojedynczego alikwotu.
Analizy sedymentologiczne i mikrosedymentologiczne dostarczają wskazówek co do pochodzenia ziaren skalenia. Cienkie przekroje ujawniają bardzo niski procent ziaren skalenia w matrycy, co zostało potwierdzone analizą samej brekcji, która zawiera minimalną ilość granitowych otoczaków. Jednak kilka znalezionych granitowych otoczaków było silnie zwietrzałych i mogło być pozakrasowym źródłem ziaren skalenia potasowego znalezionych w próbkach. Wyjaśnia to również niską wrażliwość ziaren na luminescencję. Jeśli chodzi o potencjał wybielania ziaren – pozakrasowe pochodzenie dobrze wróży wybielaniu, ale zależy to od tego, kiedy otoczaki zostały zwietrzałe i uwolniły ziarna skalenia potasowego do matrycy. Jeśli są zwietrzałe in situ, szanse na wybielanie są zmniejszone. To, w połączeniu z gwałtownym zalaniem w środowisku krasowym, nie jest najlepszym scenariuszem dla wybielania ziaren. W tej sytuacji podejście jednoziarniste jest w pełni uzasadnione, aby zidentyfikować ziarna, które są najbardziej wybielone, jednak istnieje szansa, że żadne z ziaren nie zostało wybielone. W tej sytuacji nawet pozornie dobrze wybielone ziarna mogą nadal przeceniać wiek pochówku, co oznacza, że wszystkie ziarna mają skutecznie maksymalny wiek. Jednak gdy weźmiemy pod uwagę związek między osadami a skamieniałościami, jasne jest, że osadzanie się osadów jest ostatnim zdarzeniem, które miało miejsce, ponieważ wody powodziowe zebrałyby skamieniałości z krajobrazu (lub wchłonęłyby już istniejące skamieniałości z krasowego środowiska), co spowodowałoby, że wiek osadów byłby najmłodszy — nawet jeśli jest maksymalny. Wiek osadów nie ma wpływu na wiek skamieniałości, które są wszystkie konsekwentnie starsze i wszystkie zostały nagromadzone co najmniej 104 tys. lat temu (wiek nadległego nacieku).
Uderzającą różnicę między środowiskową mocą dawki promieniowania LCC1 (3,6 Gy/kyr) i LCC2 (5,0 Gy/kyr) w tej samej jednostce brekcji można wyjaśnić heterogeniczną naturą złoża. Odwrotne uziarnienie sugeruje szybkie pojedyncze wydarzenie krasowe o rosnącej energii. Wody powodziowe osadzały gliny arenitowe przed osadzeniem klastów w gruboziarnistej, wznoszącej się sekwencji. W punkcie LCC1 stosunek klastów do matrycy wynosił ~50% i był zdominowany przez klasty wapienia mikrytowego i spartytowego; wzrósł on do ~80% w LCC2 i był zdominowany przez otoczaki granitowe, kwarcytowe i kwarcowe żyłowe pochodzące z geologii podłoża w promieniu ~20 km od jaskini. Ta duża różnica w procencie klastów i składzie wystarcza, aby wyjaśnić różnicę w środowiskowych mocach dawki promieniowania. Jednak aby upewnić się, że wartości są rzeczywiście dokładne, zastosowaliśmy spektrometrię gamma o wysokiej rozdzielczości do obu bloków. Dało to taki sam wynik dla LCC1 i nieznacznie wyższą dawkę dla LCC2 (patrz tabela uzupełniająca 3 ).
Wykorzystaliśmy dane spektrometrii gamma o wysokiej rozdzielczości z próbek LCC1 i LCC2, aby zbadać różnice w dozymetrii i sprawdzić, czy osad znajduje się w równowadze wiekowej. Spektrometria gamma o wysokiej rozdzielczości osadów jaskiniowych jest szczególnie przydatna, ponieważ dostarcza informacji o całych łańcuchach rozpadu 238 U i 232 Th; dlatego też można zbadać stężenia nuklidów potomnych, jak i macierzystych. W tym środowisku sedymentacyjnym zidentyfikowano trzy potencjalne stany nierównowagi: 1) w łańcuchu 238 U niedobór 16% 226 Ra w porównaniu do 238 U; 2) w 210 Pb niedobór od 17 do 22% w porównaniu do 226 Ra; i 3) w łańcuchu 232 Th niewielki nadmiar 7% 228 Th w porównaniu do 228 Ra. Ta nierównowaga jest bardziej widoczna w próbce LCC2 niż w osadach wokół LCC1. Jest to kolejny przykład różnic między tymi obszarami, które należy ponownie przypisać różnicom w klastach i składzie oraz zdolności osadów bardziej podtrzymywanych przez klasty w LCC2 do umożliwiania przesiąkania wody, wypłukiwania bardziej mobilnych radionuklidów, takich jak uran, i utraty radonu. Uważa się, że tor jest nieruchomy w większości osadów86 , a krótkie okresy półtrwania (t) nuklidów potomnych ( 228 Ra = 5,7 roku i 228 Th = 1,91 roku) i 210 Pb (22 lata) oznaczają, że ten efekt nadmiarów w tych radionuklidach jest nieistotny. Wypłukiwanie uranu wydaje się być większym problemem w LCC2 niż LCC1, ale niedobór 16% miałby tylko niewielki wpływ na całkowitą moc dawki i został uwzględniony w oszacowaniu błędu dla tej próbki. Obie próbki wykazują utratę radonu reprezentowaną przez niedobór 210 Pb w porównaniu do 226 Ra, ale utrata jest o 5,45% większa w LCC2. Można to również przypisać obecności większych klastów, które umożliwiają utratę radonu przez jednostkę bardziej podpartą matrycą. Jednak utrata radonu nie wyjaśnia, dlaczego LCC2 ma znacznie wyższą moc dawki, ponieważ większa utrata radonu w tej próbce powinna zamiast tego spowodować spadek mocy dawki. Klasty wapienia mogą być również źródłem nierównowagi, która jest bardziej obfita w LCC2. Ponieważ zespół mineralogiczny frakcji iłu nie zmienia się między próbkami, zmienność niedoboru 226 Ra może odzwierciedlać proporcję klastów wapienia w warstwach osadowych. Pomimo wszystkich tych różnic, główną i największą różnicą w mocy dawki środowiskowej pomiędzy próbkami jest prawie dwukrotne zwiększenie o 40K w LCC2. Może to wynikać z wyższej zawartości skalenia potasowego w zwietrzałych otoczakach granitowych.
Dawka promieniowania w środowisku brekcji (3,6–5,0 Gy/kyr) jest znacznie wyższa niż w dolnej jednostce mulistej, wynoszącej 2,7 Gy/kyr. W związku z tym uzyskane wartości D e brekcji były bardzo wysokie i wynosiły od 400 do 2000 Gy, a wartości D 0 od 600 do 1000 Gy. Oznaczało to, że niektóre ziarna były bliskie punktu nasycenia (~1000–1200 Gy) i przesuwały granice nasycenia dawki skalenia. To sprawiło, że kryteria odrzucenia stały się niezwykle ważne, aby zapewnić, że nasycone ziarna nie zostaną uwzględnione w ziarnach zaakceptowanych. Skalenie uzyskane z zwietrzałego otoczaka stanowią najlepszy test charakterystyki nasycenia tych próbek, ponieważ nigdy nie były wybielane, więc powinny wykazywać nieskończony wiek. Początkowy test zanikania z użyciem tych ziaren wykazał ziarna, które nasycają się ~1500–2000 Gy przy D0 ~600–1000 Gy. Jest to w granicach naszej oceny D e .
Ze względu na problemy związane z wypłukiwaniem uranu w środowiskach jaskiń zdecydowaliśmy się wykorzystać dawki promieniowania środowiskowego uzyskane ze spektrometrii gamma o wysokiej rozdzielczości jako bardziej konserwatywne podejście. Korzystając z tych dawek, trzy bloki wygenerowały szacunki wieku, które są zgodne z ich lokalizacją stratygraficzną i mieszczą się w ich marginesach błędu (w granicach 1 σ). Dwie szacunki wieku brekcji (LCC1 143 ± 24 i LCC2 133 ± 19 tys. lat) są statystycznie nieodróżnialne od siebie, pomimo dużej różnicy w dawkach promieniowania (odpowiednio 3,6 i 5,0 Gy/kyr). Sugeruje to, że matryca brekcji została rzeczywiście szybko zdeponowana w obrębie jednej jednostki, jak sugerują analizy sedymentologiczne. Podstawowa jednostka mulista (LCC3 248 ± 31 tys. lat) z minimalną liczbą klastów miała znacznie niższą dawkę i znacznie niższą akceptację ziarna skalenia (0,2%), co wskazuje na inny skład i/lub inne okoliczności depozycyjne. Wszystkie trzy próbki zawierały szeroki zakres wartości D e z wartościami nadmiernej dyspersji wynoszącymi od 23 do 38%. Uzasadniało to użycie MAM, który zidentyfikował ziarna, które były najbardziej wybielone przed pochówkiem w jaskini Tam Ngu Hao.
Datowanie metodą uranową nacieków
Datowanie metodą U-series nadległego nacieku w Tam Ngu Hao 2 pozwala określić minimalny wiek osadzania brekcji i towarzyszących skamieniałości. Cztery oddzielne podpróbki wywiercono ze świeżego przekroju ręcznego okazu nacieku in situ przy użyciu wiertarki ręcznej. Kalcyt pobrano z nacieku in situ przy użyciu młotka i dłuta. Sproszkowane podpróbki poddano obróbce chemicznej i pomiarom izotopowym za pomocą spektrometrii masowej 87 . Datowanie metodą U-series próbek speleothem przeprowadzono w Radioogenic Isotope Facility (The University of Queensland) przy użyciu wielokolektorowego spektrometru masowego sprzężonego indukcyjnie (MC-ICP-MS) Nu Plasma. Procedury analityczne były zgodne z poprzednimi publikacjami dla MC-ICP-MS 87 , 88 , 89 . Wiek 230 Th/ 234 U obliczono przy użyciu programu Isoplot EX 3,75 90 , a okresy półtrwania wynoszą odpowiednio 75 690 lat ( 230 Th) i 245 250 lat ( 234 U) 91 .
Komentarze do datowania metodą uranową nacieku
Nadległy naciek składa się z kalcytu niskiej jakości i ma wysoki składnik detrytyczny we wszystkich czterech podpróbkach, co odzwierciedlają wysokie poziomy stężeń 232 Th i bardzo niskie wskaźniki aktywności 230 Th/ 232 Th (0,84 do 1,0), które dały niedokładne i niewiarygodne wyniki wieku. Ten naciek ma co najmniej mniej niż 153 ± 1 tys. lat, co jest najmłodszym niekorygowanym wiekiem spośród wszystkich czterech podpróbek (tj. maksymalny wiek podpróbki). Nieradiogeniczna poprawka 230 Th przy użyciu standardowej wartości aktywności Bulk-Earth 230 Th/ 232 Th wynoszącej 0,825 z założoną niepewnością 50% dla nieradiogennej poprawki 230 Th spowodowała bardzo duże powiększenia błędów, przy czym jedna z czterech podprób zwróciła ujemną wartość wieku, co wyraźnie wskazuje na nadmierną korektę nieradiogennego 230 Th. Lepsze oszacowanie najbardziej wiarygodnej poprawki opiera się na założeniu, że rzeczywisty współczynnik aktywności detrytycznej 230 Th/ 232 Th powinien być niższy niż wartość bulk-Earth wynosząca 0,825. Aby uzyskać lepsze oszacowanie, nanosimy zmierzone współczynniki aktywności 234 U/ 232 Th na zmierzone współczynniki aktywności 230 Th/ 232 Th, co definiuje najlepiej dopasowaną linię z wartością przecięcia 0,5 na osi 230 Th/ 232 Th. Innymi słowy, ten przecięcie reprezentuje wspólny początkowy (lub detrytyczny) stosunek aktywności 230 Th/ 232 Th czterech podpróbek. Tak więc nowy schemat korekcji wykorzystujący nieradiogenną wartość aktywności 230 Th/ 232 Th wynoszącą 0,5 jest bardziej odpowiedni. Na podstawie tego nowego schematu cztery podpróbki dają nierozróżnialne skorygowane wieki ze średnią ważoną 104 ± 27 tys. lat (szczegóły w tabeli uzupełniającej 4 ). Podczas próby ustalenia wieku leżącej pod spodem brekcji, ten wynik jako samodzielny szacunek wieku nie jest szczególnie przydatny, ale jako część sekcji chronologicznej zapewnia „minimalny” szacunek wieku dla całej sekwencji stratygraficznej nacieku, brekcji i leżącej pod spodem jednostki mułowej i dlatego został wykorzystany w modelu bayesowskim.
Sprzężony szereg uranowy i rezonans spinowy elektronów (US-ESR)
Trzy zęby wołowe (TNH2-10/CC10, TNH2-11/CC11, TNH2-12/CC12) odzyskane z górnej brekcji zawierającej skamieniałości (LU2) zostały użyte do bezpośredniego datowania metodą sprzężonej US-ESR zgodnie z protokołem Joannes-Boyau i in. 92 , 93 , 94 (ryc. 1b ). Wszystkie trzy zęby zostały najpierw przecięte na pół za pomocą precyzyjnej piły diamentowej, a następnie wypolerowane do gładkości 50 μm, aby odsłonić płaską powierzchnię zębiny i szkliwa do przesiewowego badania stężenia 238 U przy użyciu ESI NW213 i ICPMS kwadrupol Agilent 7700 do stężenia. Następnie mały fragment zębiny i szkliwa z każdego zęba fauny został usunięty za pomocą 300 μm grubości diamentowego ostrza przymocowanego do ręcznej piły obrotowej do analiz serii U przy użyciu LA-MC-ICPMS Thermo Neptune plus sprzężonego z jednostką ablacji laserowej ESI NW193 (Rys. uzupełniający 11 ). Stosunek izotopów i stężenia uzyskano odpowiednio za pomocą analiz LA-MC-ICPMS i kwadrupolowego ICPMS. Średnia wartość została obliczona dla każdej tkanki zębowej i wykorzystana w modelu US-ESR. Linię bazową i dryf skorygowano za pomocą szklanego dysku NIST 612, podczas gdy skamieniały ząb hipopotama o znanym stężeniu serii U został użyty do skorygowania stosunków 234 U/ 238 U i 230 U/ 238 U oraz oceny dokładności pomiarów. Inny fragment szkliwa z każdego zęba został użyty do pomiarów ESR. Datowanie ESR przeprowadzono przy użyciu spektrometru pasma X Freiberg MS5000 przy amplitudzie modulacji 1 G, mocy 2 mW, przemiataniu 100 G i częstotliwości modulacji 100 kHz, sprzężonego z komorą napromieniowania rentgenowskiego Freiberg, która zawiera działo rentgenowskie Varian VF50 o napięciu 40 kV i natężeniu prądu 0,5 mA 95 , 96 . Procedury dopasowania przeprowadzono przy użyciu oprogramowania MCDoseE 2.0, które wykorzystuje podejście oparte na schemacie bayesowskim, gdzie rozwiązaniem jest pełny rozkład prawdopodobieństwa równoważnika dawki 97 .
Fragmenty szkliwa CC10, CC11 i CC12 pobrane z każdej próbki i użyte do pomiarów ESR miały bezpośredni kontakt z zębiną po jednej stronie i bezpośredni kontakt z osadem po drugiej stronie (bez cementu). Zewnętrzna powierzchnia szkliwa (mająca kontakt z otaczającym osadem) i zębina bezpośrednio do niej przytwierdzona zostały usunięte za pomocą obrotowego narzędzia z ostrzem diamentowym. Jednocześnie usunięto 100 μm z każdej strony, aby uniknąć udziału cząstek alfa. Każdy fragment zęba został następnie pocięty na trzy oddzielne części (A, B i C), przy czym A (50 Gy) i B (200 Gy) zostały wstępnie napromieniowane jedną dawką promieniowania gamma przed napromieniowaniem rentgenowskim, a C napromieniowano wyłącznie promieniami rentgenowskimi. Fragmenty zamontowano na teflonowym uchwycie próbki, umożliwiając bezpośrednie wystawienie fragmentu na działanie źródła promieni rentgenowskich za 200 μm osłoną aluminiową. Aby oszacować równoważną dawkę ESR (D e ), każdy fragment napromieniowano siedem razy, stosując wykładniczo zwiększające się czasy napromieniowania (tj. 90 s, 390 s, 975 s, 1891 s, 3604 s, 7380 s i 12382 s ze średnią mocą dawki dla CC10, CC11 i CC12 wynoszącą odpowiednio 0,231 Gy/s, 0,203 Gy/s i 0,249 Gy/s). Emisję promieniowania rentgenowskiego otrzymaną przez zęby bydlęce skalibrowano przy użyciu dodatkowych znanych dawek promieniowania gamma na fragmentach A i B wykonanych w Australijskiej Organizacji Nauki i Technologii Jądrowej (ANSTO). Podczas każdego etapu napromieniowania rejestrowano wyjście pistoletu rentgenowskiego, aby umożliwić dokładne określenie dawki otrzymanej przez próbkę na każdym etapie. Krzywe zależności dawka-odpowiedź (DRC) uzyskano przez uśrednienie szczytowych natężeń T 1 -B 2 ESR zarejestrowanych dla każdej dawki promieniowania w pomiarach 180° (krok 20 ° ) i połączenie ich w jedno widmo 98 . Składowe izotropowe i poprawki bazowe zastosowano równomiernie w zmierzonych widmach 94 .
Podjęto próbę bezpośredniego datowania TNH2-1, ale aby zminimalizować uszkodzenia cennej próbki, wykorzystano powierzchowną warstwę zębiny resztkowej na szczątkach korzenia. Niestety analizy wykazały wysoki stopień diagenezy mikropróbki, co całkowicie przyćmiło jakąkolwiek sensowną interpretację wieku skamieniałości.
Komentarze dotyczące sprzężonych szeregów uranowych i rezonansu spinowego elektronów (US-ESR)
Niezorientowane komponenty (NOCOR) sygnału ESR wynoszące odpowiednio 16%, 14 i 8% dla TNH2-10/CC10, TNH2-11/CC11 i TNH2-12/CC12 zostały wyekstrahowane z sygnału przy użyciu metody Joannes-Boyau 94. Końcowe wartości D e uzyskano, dopasowując pojedynczy wykładniczy rozkład nasycenia (SSE) do intensywności ESR i wybierając odpowiednią maksymalną dawkę napromieniowania (Dmax), aby uniknąć niedokładności oszacowania dawki 99 . Wszystkie błędy obliczono z interwałem 2-sigma. Osady otaczające zęby zebrano i przeanalizowano w roztworze za pomocą ICPMS po metodzie trawienia kwasem (1:3 azot i HCL). Zewnętrzną dawkę promieniowania gamma obliczono, używając zawartości osadu zmierzonej za pomocą ICPMS i zakładając pełną geometrię 4π. Całkowite moce dawek dla TNH2-10/CC10, TNH2-11/CC11 i TNH2-12/CC12 obliczono na poziomie 1482 ± 224, 1253 ± 192 i 1187 ± 208, co daje wiek US-ESR wynoszący odpowiednio 164 ± 24 tys. lat, 149 ± 22 tys. lat i 140 ± 23 tys. lat, stosując metodę Monte Carlo ( Tabela uzupełniająca 2 ) i zakładając równowagę dla reszty łańcucha rozpadu. Jednakże dyfuzja uranu pasuje do modelu układu otwartego z akumulacją uranu na połączeniu szkliwa z zębiną i w pęknięciach, a także obecnością wypłukiwania uranu i wzbogacenia toru 232. W przypadku próbki TNH2-10/CC10 należało założyć stosunek 230 Th/ 234 U w szkliwie (patrz tabela uzupełniająca 2 (*)) z powodu wypłukiwania i/lub wzbogacenia osadu (zmierzony stosunek 230 Th/ 234 U >1, a stężenie U jest niskie, podczas gdy tor jest wysokie). Niestety, w celu modelowania wieku przy użyciu obliczeń Monte Carlo z Shao i in. 100 , użyliśmy tego samego stosunku 230 Th/ 234 U do stosunku zębiny dla szkliwa, wprowadzając znaczące założenie do obliczeń wieku tej konkretnej próbki. W tych warunkach łączna ocena wieku umieszcza sekwencję nieco starszą niż 150 tys. lat. TNH2-10/CC10 pozostaje nieco starszy, ale mieści się w granicach błędu TNH2-11/CC11 i TNH2-12/CC12, a zatem jest statystycznie nieodróżnialny. Ponadto obliczono , że równoważnik dawki (D e ) TNH2-10/CC10 jest 1,3 i 1,5 razy większy niż dla TNH2-11/CC11 i TNH2-12/CC12 (Tabela uzupełniająca 2 ). Ponadto stężenie NOCOR dla TNH2-12/CC12 mogło zostać niedoszacowane z powodu niezwykle słabej odpowiedzi kątowej fragmentu. Podobna zawartość NOCOR jak dla TNH2-10/CC10 lub TNH2-11/CC11 zwiększyłaby D eo ~8%, co przybliżyłoby wiek do TNH2-11/CC11. Przestrzenny rozkład wyników serii U dla każdego zęba pozwala lepiej zrozumieć proces dyfuzji w tkankach zęba (patrz Rycina uzupełniająca 12a–c ). Poza punktową intruzją osadów w pęknięciach i innych ubytkach zębów (Rycina uzupełniająca 12c ), rozkład jest charakterystyczny dla ścieżki dyfuzji z wychwytem z jamy miazgi do zębiny lub w mniejszym stopniu z otaczających osadów do zewnętrznej warstwy szkliwa w kontakcie z osadami. Wiek i gradient stężenia obserwowany wraz z odległością od początku wychwytu nie znajduje odzwierciedlenia w zmierzonym stosunku serii U. Tak więc potencjalne wypłukiwanie i/lub późne wzbogacenie mogło mieć miejsce, ale byłoby marginalne w odniesieniu do obliczenia wieku. Ten wzór dyfuzji wzmacnia wyniki wieku serii U i daje średnią ocenę wieku wynoszącą 189 ± 9 tys. lat, 132 ± 13 tys. lat i 124 ± 16 tys. lat (2-sigma) odpowiednio dla TNH2-10/CC10, TNH2-11/CC11 i TNH2-12/CC12. Wartość p (funkcja dyfuzji uranu w tkankach zębowych) we wszystkich trzech zębach zbliża się do −1 lub innymi słowy, jest równoważna wczesnemu wychwytowi izotopów przez zęby. To z kolei wyjaśnia, dlaczego wyniki datowania serii U i US-ESR mieszczą się w granicach błędu. Podsumowując, wyniki US-ESR zębów fauny dają średni ważony wiek szczątków ludzkich wynoszący 151+/− 37 tys. lat. Wreszcie, podczas gdy skamieniałe zęby zostały odkopane na głębokości 20 cm od powierzchni, struktura brekcji wskazuje, że duża część uległa erozji w niedawnym czasie. Dlatego też moc dawki zewnętrznej została zmodelowana przy założeniu pełnej 30-centymetrowej kuli brekcji otaczającej każdą próbkę fauny. Jeśli zmodelujemy moc dawki zewnętrznej do obecnej konfiguracji pochówku, moc dawki zewnętrznej wyniesie 699+/−121 mGy/kyr. Ze względu na znaczącą rolę składowej zewnętrznej w całkowitej mocy dawki, szacunki wieku zostałyby przesunięte do 190+/−28 kyr, 177+/−26 kyr i 168+/−27 kyr odpowiednio dla CC10, CC11 i CC12.
Modelowanie jednostki brekcji i towarzyszących jej skamieniałości
Aby ocenić niepewności zintegrowanego podejścia do datowania stanowiska (tabele uzupełniające 1–4 ) , przeprowadzono modelowanie bayesowskie dla wszystkich niezależnych szacunków wieku przy użyciu oprogramowania OxCal (wersja 4.2) 101 dostępnego pod adresem https://c14.arch.ox.ac.uk/oxcal.html (rys. uzupełniający 3). Analizy uwzględniły rozkłady prawdopodobieństwa poszczególnych wieków, ograniczenia narzucone przez relacje stratygraficzne oraz zgłoszoną minimalną lub maksymalną naturę niektórych indywidualnych szacunków wieku. Każdy indywidualny wiek uwzględniono jako rozkład Gaussa ( ze średnią i odchyleniem standardowym zdefiniowanymi przez szacunek wieku i ich powiązane niepewności), a wynikowe zakresy wieku dla każdej jednostki przedstawiono przy 1 σ.
Nie podjęto próby usunięcia zidentyfikowanych wartości odstających. Wynika to z faktu, że nie znamy podstawowego „prawdziwego” modelu głębokości wieku i stosujemy kilka różnych metod datowania, więc trudno jest określić kryterium identyfikacji prawdziwych wartości odstających. Zamiast tego podejścia wyraźnie określiliśmy minimalny i maksymalny wiek, gdzie było to właściwe, aby to zrobić zgodnie z charakterem metod datowania i jakością wyników. Uważamy, że jest to lepsza metoda w porównaniu z analizą wartości odstających w tym kontekście, ponieważ unika ona niepotrzebnych uprzedzeń (np. w wyborze kryterium) i reprezentuje bardziej konserwatywne podejście.
Komentarze dotyczące modelowania jednostki brekcji i towarzyszących jej skamieniałości
Szacunki wieku są równowiekowe, a niepewności są stosunkowo niewielkie. Jako takie, zidentyfikowane wieki graniczne nie są wrażliwe na usunięcie poszczególnych dat lub na zmiany, na przykład, rozdzielczości obliczeń modelu. Żadna ze zmian, które wprowadziliśmy do konfiguracji modelu, nie spowodowała zauważalnych różnic w wynikach modelu wieku. Wiek brekcji w Tam Ngu Hao 2 został ostrożnie oszacowany jako granica między leżącą wyżej jednostką naciekową (104 ± 27 tys. lat) a szacunkami wieku brekcji i skamieniałości z wnętrza brekcji (131 ± 15 tys. lat) oraz granicą między brekcją a leżącą niżej jednostką mułową (164 ± 17 tys. lat), uwzględniając wszystkie ograniczenia opisane powyżej i wynikające z nich szacunki wieku (tabele uzupełniające 1–4 ) . Ostateczne modelowane wieki przedstawiono na rysunku uzupełniającym 3 , a skrypt przedstawiono na rysunku uzupełniającym 13 . Szacunki wieku brekcji, pochodzące zarówno z datowania zębów metodą US-ESR, jak i datowania macierzy brekcji metodą pIR-IRSL, są współczesne i mieszczą się w granicach błędu, pomimo ograniczenia dwóch różnych zdarzeń, tj. odpowiednio śmierci fauny i osadzania jednostki brekcji. Wskazuje to, że czas między śmiercią a ostatecznym pochówkiem w jaskini był bardzo krótki, a brekcja została szybko osadzona jako jedna jednostka.
Szczegółowa analiza morfologiczna zęba
Opis TNH2-1
TNH2-1 to izolowany lewy stały zarodek korony zęba trzonowego żuchwy, którego wzrost korzenia został właśnie zainicjowany i osiągnął ~1,8 mm poniżej korony (ryc. 2a–f ). Szkliwo jest dobrze zachowane, z błyszczącą beżową patyną z bielszymi plamami. Widoczna jest niewielka hipoplazja szkliwa w kierunku szyjki macicy i drobne wgłębienia na środkowej wysokości korony na powierzchniach policzkowej i mezjalnej, ale nie ma zmian próchnicowych. Zębina jest ciemnobrązowa na powierzchni zewnętrznej z kilkoma miejscowo zmienionymi obszarami, co pokazuje, że zębina pod nią jest bardziej żółta. Zarys okluzyjny jest subpentagonalny. Brak zużycia okluzyjnego (stadium 1 102 ) i międzyzębowych powierzchni mezjalnych i dystalnych wskazuje, że ząb nie wyrósł w chwili zgonu. Istnieje pięć głównych guzków (protokonid>entokonid≥hypokonid>metakonid>hypokonid) łączących się w + rowku na środku zagłębienia zwarciowego, a także tuberculum sextum (C6) zlokalizowane na dystalnym brzeżnym grzebieniu hipokonidu (Ryc. 3 ). Na mezjalnym grzebieniu hipokonidu znajduje się wyraźne ramię, które objawia się jako mały guzek na powierzchni szkliwa. Istnieje wysoki i ciągły środkowy środkowy grzebień trygonidu (z niewielkim zagłębieniem centralnie) i przerwany dystalny grzebień trygonidu z segmentem policzkowym rozpoczynającym się na dystalnym grzbiecie protokonidu i segmentem językowym rozpoczynającym się na dystalnym grzbiecie metakonidu (typ A 103 ). Ten wzór grzebienia trygonidowego odpowiada wzorcowi typu 8 klasyfikacji Martineza de Pinillosa i współpracowników 103 , z ciągłym grzebieniem środkowotrygonidowym i nieciągłym dystalnym grzebieniem trygonidowym powstającym z dystalnych segmentów. Istnieją również krótsze grzebienie ustawione dystalnie do dystalnego grzebienia trygonidowego i większe grzebienie biegnące od guzków talonidowych w dół w kierunku środka zagłębienia zwarciowego. Przednia dołka jest wąska, rozciągnięta policzkowo-językowo i głęboka (typ 4 według ASUDAS 104 , 105 ). Nie ma protostylidu (typ 0 według ASUDAS 104 , 105 ), chociaż wykryto słabą półkę policzkową wzdłuż powierzchni policzkowej w EDJ (ryc. 3b ). W EDJ można zaobserwować, że mezjalne rogi zębiny są bardziej centralnie umieszczone i bardziej pochylone w kierunku środka zagłębienia okluzyjnego niż dystalne. W widoku od dołu można dostrzec pięć rogów miazgi, pomimo niedojrzałego stadium komory miazgi.
Całkowicie pięciokątny kształt korony z niemal równoległymi ścianami policzkową i językową, obecność kompletnego środkowego grzebienia trygonicznego, wyraźnie uwypuklony hypoconulid i C6, a także średnica wydłużona w kierunku mezjodystalnym (patrz ryc. 2 i 3 ; ryc. uzupełniająca 2 ; tabele uzupełniające 10 i 11 ) sugerują, że jest to raczej pierwszy lub drugi ząb trzonowy, a nie trzeci ząb trzonowy. Analizy statystyczne przeprowadzone na kształcie EDJ (patrz Tabela uzupełniająca 16 ) klasyfikują TNH2-1 jako M2, ale jego wyraźne podobieństwo do M1 żuchwy Xiahe (co potwierdza bliskie położenie obu zębów w analizach GM EDJ) i oczywista różnica w stosunku do M2 tej drugiej, która wykazuje nietypową morfologię zwarcia (z wyraźnym przedłużeniem policzkowo-językowym zagłębienia zwarcia związanego z obecnością nadliczbowego korzenia poniżej; patrz ref. 15 ) rzucają wątpliwości na jego pozycję metameryczną (patrz Rys. uzupełniający 5 ). Fakt, że M1 i M2 okazów Xiahe są klasyfikowane jako M1 (w analizach semilandmark; Tabela uzupełniająca 16 ) lub jako M2 (w analizach DSM; Tabela uzupełniająca 16 ) może również wskazywać na szczególną cechę zębów trzonowych Denisovana. Rzeczywiście, podczas gdy niepewne przypisanie M2 nie jest zaskakujące, biorąc pod uwagę wyżej wspomniany nietypowy kształt EDJ, M1 nie wykazuje żadnej anomalii i powinno być klasyfikowane jako M1 w obu analizach. Jak wskazuje trudność w identyfikacji zębów trzonowych Denisova 4 i 8 jako M2 lub M3, ten niejednoznaczny wynik dotyczący zębów trzonowych Xiahe może sugerować, że zmienność metameryczna zębów trzonowych Denisova różni się od zmienności grup uwzględnionych tutaj jako odniesienie, uniemożliwiając nam jednoznaczne przypisanie TNH2-1 do M1 lub M2, jeśli należy do tej drugiej grupy.
Analizy morfologiczne
Mikrotomografia rentgenowska
Próbkę TNH2-1 zeskanowano przy użyciu instrumentu X-ray microfocus (X-µCT) diondo d3 w Instytucie Antropologii Ewolucyjnej Maxa Plancka w Lipsku w Niemczech. Akwizycje przeprowadzono zgodnie z następującymi parametrami: 130 kV, 70 µA, filtr mosiężny 0,5 mm, 2970 obrazów wykonanych w zakresie 360° (~0,12° kroku kątowego) ze średnią 2 (uśrednianie klatek), czas integracji 1000 ms. Ostateczne objętości zrekonstruowano przy rozmiarze woksela 9,14 µm. Akwizycje mikrotomograficzne porównawczych okazów kopalnych i żyjących hominidów przeprowadzono przy użyciu różnych urządzeń, w tym X-µCT i promieniowania synchrotronowego (SRX-µCT) i zrekonstruowano przy rozmiarach wokseli od 10,25 do 57,50 µm.
Przetwarzanie danych
Wykonano półautomatyczną segmentację opartą na progach w programie Avizo 8.0 (FEI Visualization Sciences Group), stosując metodę połowy maksymalnej wysokości (HMH 106 ) i protokół progowania obszaru zainteresowania (ROI-Tb 107 ), dokonując wielokrotnych pomiarów na różnych wycinkach wirtualnego stosu 108 . Następnie dla każdego okazu wygenerowano rekonstrukcję objętościową.
Wymiary korony
Średnice korony zęba w części mezjodystalnej i policzkowo-językowej TNH2-1 mierzono na oryginalnym okazie za pomocą suwmiarki Mitutoyo Digimatic (z dokładnością do 0,1 mm), a następnie na wirtualnej powierzchni wygenerowanej po segmentacji zapisu mikrotomograficznego. Pomiary te porównano ze średnicami korony zęba H. erectus sl (HE), H. antecessor (HA), europejskiego Homo środkowoplejstoceńskiego (EMPH), azjatyckiego Homo środkowoplejstoceńskiego (AMPH), neandertalczyków (NEA), górnopaleolitu H. sapiens (UPMH) i holoceńskich ludzi (HH) (patrz tabele uzupełniające 9 , 10 ). Skorygowane analizy Z-score109 , 110 przeprowadzono na średnicach mezjodystalnych i policzkowo-językowych TNH2-1 w porównaniu z porównawczymi okazami/próbkami. Ta metoda statystyczna umożliwia porównanie niezrównoważonych i małych próbek, co często ma miejsce w przypadku zapisu kopalnego, wykorzystując odwrotny rozkład t-Studenta według wzoru: [(xm)/(s*sqrt(1 + 1/n)]/(Student.t.inverse(0,05;n-1)), gdzie x jest wartością zmiennej; m jest średnią tej samej zmiennej dla próbki porównawczej; n jest liczebnością próbki porównawczej; s jest odchyleniem standardowym próbki porównawczej (Tabela uzupełniająca 11 ).
Proporcje tkanki korony 3D
Trzy zmienne opisujące proporcje tkanki korony zęba mierzono cyfrowo na TNH2-1: procent objętości korony, który stanowi zębina i miazga (Vcdp/Vc; %), średnia grubość szkliwa (3D AET; mm) i bezłuskowa względna grubość szkliwa 3D (3D RET) 111 , 112 , 113 . Wewnątrzobserwacyjne i międzyobserwacyjne testy dokładności pomiarów przeprowadzonych przez dwóch obserwatorów wykazały różnice <5%. Przeprowadzono skorygowane analizy Z-score 109 , 110 na parametrach proporcji tkanki korony zęba dla TNH2-1 w porównaniu z porównawczymi okazami/próbkami.
Geometryczna analiza morfometryczna kształtu połączenia szkliwa z zębiną (EDJ)
Zastosowaliśmy podejście dopasowania powierzchni dyfeomorficznej (DSM) do analizy konformacji EDJ. Ta metoda konstruuje atlasy kształtów przy użyciu prądów matematycznych (funkcji do analizy rozmaitości), w których zachowana jest topologia szablonu, a parametry deformacji są optymalizowane niezależnie od parametrów kształtu 114 , 115 , 116 ). Użyliśmy oprogramowania Deformetrica v. 4.3 ( https://www.deformetrica.org ) do wygenerowania globalnego średniego kształtu z zestawem dyfeomorfizmów odnoszących globalny średni kształt do każdego osobnika i danych wyjściowych (punkty kontrolne i pędy deformacji) do wykonania analiz statystycznych oraz zbadania zmienności kształtu EDJ i klasyfikowania danych. To podejście zostało już wykorzystane do zbadania kształtu EDJ kopalnych homininów i okazało się niezawodną metodą rozróżniania taksonów 117 , 118 . Dopasowane modele oparte na deformacji zastosowano do kompletnego EDJ zdziesiątkowanego do 50 000 wielokątów i wyrównanego przez superpozycję Prokrusta. Gładkie i odwracalne deformacje połączone z metryką prądów, która definiuje odległość między powierzchniami EDJ, pozwoliły nam określić stopień zmienności kształtu między badanymi okazami. Następnie globalny średni kształt i funkcje deformacji zostały wykorzystane do klasyfikacji danych. Porównaliśmy również wyniki analiz DSM z wynikami uzyskanymi za pomocą klasycznej geometrycznej metody morfometrycznej opartej na punktach orientacyjnych i półpunktach orientacyjnych umieszczonych wzdłuż okluzyjnych i szyjkowych brzegów EDJ. Dokładniej rzecz biorąc, cztery anatomiczne punkty orientacyjne umieszczono na czubku rogu zębiny czterech guzków pierwotnych (protokonid, metakonid, entokonid i hypokonid), a pierwszy zestaw półpunktów orientacyjnych umieszczono wzdłuż górnej części grzbietu brzeżnego łączącego rogi zębiny (zaczynając od góry protokonidu i przesuwając się w kierunku językowym), podczas gdy drugi zestaw półpunktów orientacyjnych umieszczono wzdłuż szyjki macicy, zaczynając poniżej rogu zębiny protokonidu (konkretnie, mezjalno-policzkowego rogu szyjki macicy) i przesuwając się w kierunku mezjalnym 39 . W przypadku braku małych części szyjki macicy szacowano jej położenie. W większości przypadków punkty orientacyjne szyjki macicy umieszczono na modelu powierzchni całej korony zębiny, ponieważ jest to najbardziej odpowiednie i ułatwia umieszczanie punktów orientacyjnych na próbkach, w których brak szkliwa wpływa na kompletność powierzchni EDJ. Geometrycznie odpowiadające sobie półpunkty orientacyjne 119 wyprowadzono w programie Mathematica (Wolfram Inc.) przy użyciu procedury programowej 120 , 121 zgodnie z protokołem pomiaru opisanym szczegółowo w pracach Skinner et al. 122 , 123. Gładką krzywą interpolowano przy użyciu funkcji spline sześciennej. Spline sześcienny dopasowano, zaczynając od pierwszego punktu orientacyjnego i przesuwając się w kierunku językowym do ostatniego punktu. Interpolowane krzywe zostały następnie ponownie pobrane, aby uzyskać identyczną liczbę punktów wśród okazów. Następnie półpunkty orientacyjne grzbietu brzeżnego EDJ zostały rzutowane na krzywą, dzieląc krzywą na cztery sekcje (protokonid do metakonidu: 12 punktów orientacyjnych; metakonid do entokonidu: 12 punktów orientacyjnych; entokonid do hipokonidu: 24 punkty orientacyjne; hipokonid do protokonidu: 12 punktów orientacyjnych). W przypadku półpunktów orientacyjnych na szyjce macicy wygenerowano 30 punktów orientacyjnych rozmieszczonych w równych odstępach wzdłuż krzywej szyjki macicy.
W przypadku obu geometrycznych podejść morfometrycznych przeprowadziliśmy następnie uogólnione analizy Prokrustesa, a następnie analizy głównych składowych (PCA). Zbadaliśmy strukturę i separację grup wzdłuż wyników PC, sprawdzając, czy grupy istniały już wcześniej w PCA (Rys. uzupełniający 7 ). Zarówno podejście DSM, jak i oparte na punktach orientacyjnych wykazują znaczny stopień separacji między taksonami wzdłuż pierwszych trzech osi PC (Rys. uzupełniający 7 ). Obliczyliśmy analizy zmiennych kanonicznych (CVA) i analizy głównych składowych między grupami (bgPCA). Następujące trzy grupy zostały wykorzystane zarówno w CVA, jak i bgPCA: H. erectus sl (HE), europejscy środkowoplejstoceńscy Homo i neandertalczycy (EMPH-NEA), plejstoceński i holoceński H. sapiens (MH). TNH2-1 i dwa zęby trzonowe (M1 i M2) żuchwy Xiahe zostały następnie rzutowane a posteriori w CVA i bgPCA. Analiza CVA została przeprowadzona na podstawie kilku pierwszych głównych składowych wyjaśniających ~90% całkowitej wariancji 124 (18 w analizie opartej na punktach orientacyjnych i 24 w analizie opartej na odkształceniach; rys. 5a , rys. uzupełniający 7 ). Przeprowadziliśmy również krzyżowo walidowane analizy zmiennych kanonicznych (CV CVA) nadal oparte na kilku pierwszych głównych składowych wyjaśniających ~90% całkowitej wariancji zarówno dla danych DSM, jak i opartych na punktach orientacyjnych (rys. uzupełniający 7 ). Analiza CV CVA została przeprowadzona najpierw w celu sprawdzenia prawdopodobieństwa, że okaz TNH2-1 reprezentuje M1 lub M2 (tabela uzupełniająca 16 ), a następnie z tymi samymi grupami dla bgPCA w celu oceny przewidywania taksonomicznego (tabela uzupełniająca 17 ). Analiza bgPCA została obliczona na podstawie momentów odkształcenia (rys. 5b ) i reszt Prokrustesa (rys. uzupełniający 7 ). Analizy DSM i punktów orientacyjnych dały podobne wyniki dla CVA i bgPCA, chociaż metoda DSM lepiej rozróżnia grupy (rys. 5b , rys. uzupełniający 7 ). Fakt, że dwa różne podejścia GM (DSM i punkty orientacyjne) odtwarzają porównywalną dyskryminację grup i rozkład przy użyciu CVA lub bgPCA, które są obliczane przy użyciu różnych danych wejściowych (odpowiednio wybranego zestawu wyników PC i momentów/reszt Prokrustesa), sugeruje, że wyniki geometrycznych analiz morfometrycznych są solidne. Postępowaliśmy również zgodnie z zaleceniami Cardiniego i Polly’ego dotyczącymi ważności bgPCA 125 , obliczając krzyżowo walidowane bgPCA (CV bgPCA; rys. uzupełniający 7 ) i porównując je z bgPCA (rys. 5b , rys. uzupełniający 7h), pokazując, że oba wykazują ten sam stopień dyskryminacji grupowej. Wyniki CV CVA (Tabela uzupełniająca 17 ) i CV bgPCA pokazują wysoki poziom poprawnej klasyfikacji (HE: 94,1%; EMPH-NEA: 98,0%; MH: 88,2% dla bgPCA na podstawie DSM z trzema grupami). Aby dalej przetestować, czy podział grup nie jest sztuczny, obliczyliśmy również bgPCA z pięcioma grupami zamiast trzech (w tym H. erectus i oddzielając środkowoplejstoceńskich i późnoplejstoceńskich neandertalczyków, a także plejstoceńskich współczesnych ludzi i holoceńskich ludzi; Rys. uzupełniający 7 ). Analizy te pokazują niemal identyczne rozróżnienia taksonomiczne niż w bgPCA na podstawie trzech grup ( H. erectus , neandertalczycy i współcześni ludzie; Rys. 5b , Rys. uzupełniający 7 ), wskazując, że podział grup odzwierciedla rzeczywistość biologiczną. Ogólnie rzecz biorąc, analizy PCA i bgPCA, CV bgPCA i CVA oparte na punktach orientacyjnych i DSM są spójne ze sobą i potwierdzają, że grupy obserwowane w bgPCA nie są fałszywe 126 . Analizy przeprowadzono przy użyciu pakietów ade4 v.1.7-6 127 i Morpho v. 2.7 128 dla R v.3.4 129 . Allometria została przetestowana w analizach półpunktów orientacyjnych przy użyciu regresji wielokrotnej 130 w której zmienną objaśniającą jest rozmiar centroidu, a zmiennymi zależnymi są wyniki PC i bgPC. We wszystkich PCA i bgPCA pierwsze dwa składniki nie wykazują allometrii ( wartość p > 0,05). Różnice między okazami reprezentują zatem zmienność kształtu.
Analiza starożytnych białek
Ekstrakcję białek szkliwa przeprowadzono zgodnie z protokołami szczegółowo opisanymi w innym miejscu 34 . Krótko mówiąc, małe próbki szkliwa demineralizowano w kwasie solnym (HCl) lub kwasie trifluorooctowym (TFA), czyszczono na końcówkach C18, a następnie analizowano przy użyciu nanoLC-MS/MS bez trawienia proteazą lub redukcji cysteiny i alkilacji.
Proteom szkliwa TNH2-1 analizowano przy użyciu wcześniej opublikowanego przepływu pracy 34 . Próbkę przetwarzano w czystym laboratorium odpowiednim do analizy starożytnego DNA i białka. Szkliwo oczyszczono mechanicznie, a dwie małe próbki (12 i 13 mg) usunięto i sproszkowano w celu ekstrakcji białka. W celu demineralizacji proszki szkliwa zawieszono w 1,5 M (5,5% w/w) HCl lub 0,88 M (10% w/w) TFA i inkubowano w temperaturze pokojowej przez noc. Peptydy ekstrahowano z supernatantu i odsolono przez ekstrakcję w fazie stałej przy użyciu C18 Stagetips 131 . Próby laboratoryjne przygotowano przy użyciu tej samej procedury. Peptydy suszono próżniowo, rekonstytuowano w 0,1% TFA 5% ACN i oznaczano ilościowo przy użyciu spektrofotometru Nanodrop przy 280 nm. Około 500 ng peptydów załadowano na kolumnę o średnicy wewnętrznej 75 µm i długości 15 cm, wypełnioną samodzielnie kulkami Reprosil-Pur C18 o średnicy 1,9 µm (Dr. Maisch) z emiterem krzemionkowym ciągnionym laserowo. Anality rozdzielono przy użyciu EASY-nLC 1200 (Thermo Scientific) i dwóch różnych gradientów przy użyciu 0,1% TFA 99,9% wody jako rozpuszczalnika A i 0,1% TFA 80% ACN 19,9% wody jako rozpuszczalnika B przy szybkości przepływu 350 nl/min. Krótszy 45-minutowy gradient wzrósł z 8 do 15% B w ciągu 25 min, do 30% B w ciągu 15 min i do 45% B w ciągu 5 min. Dłuższy 90-minutowy gradient wzrósł z 8 do 20% B w ciągu 75 min, z 20 do 30% B w ciągu 10 min i z 30 do 45% B w ciągu 5 min. LC połączono z Orbitrap Q-Exactive HF-X (Thermo Scientific) 132 , uzyskując pełne skany MS od 350 do 1400 m/z przy rozdzielczości 60 000 przy 200 m/z, celu 3e6 AGC i maks. 45 ms. Po czasie wtrysku (IT) nastąpiło 10 zależnych od danych skanów MS2 z szerokością izolacji 1,3 m/z i fragmentacją HCD przy znormalizowanej energii zderzenia 28 i zmierzonej przy rozdzielczości 45 000, celu 1e5 AGC, maks. 45 ms. IT i pierwszej masie 100 m/z. Surowe pliki analizowano za pomocą programu Maxquant w wersji 1.6.0.17 133, przeszukując wewnętrzną bazę danych zawierającą sekwencje białek oczekiwane w proteomie szkliwa Hominidae 35 , 36 . Peptydy kandydujące wygenerowano przez trawienie „niespecyficzne” i umożliwiono do 3 z następujących zmiennych modyfikacji: deamidacja (NQ), hydroksyprolina (P), fosforylacja (STY), utlenianie (MW), dioksydacja (W), tworzenie kynureniny (W) i konwersja argininy do ornityny (R), które wybrano na podstawie wcześniej opublikowanej strategii otwartego wyszukiwania 134 przy użyciu funkcji wyszukiwania „peptydów zależnych” programu Maxquant. Przeszukiwanie programu Maxquant obejmowało również proteom szkliwa człowieka (Ø1952, samiec, Homo sapiens ) sprzed około 300 lat, który został wcześniej przygotowany i przeanalizowany w identyczny sposób 35 , a także liczne ekstrakcje i puste próbki spektrometrii masowej.
Próbki ekstrakcyjne i ślepe spektrometrii masowej nie zawierały żadnych białek, co wskazuje na czyste warunki ekstrakcji i instrumentu podczas analiz. Następnie białka reprezentowane przez pojedynczy peptyd i prawdopodobne zanieczyszczenia usunięto z analiz. Pozostawiło to niewielką liczbę znanych białek szkliwa do dalszych analiz filogenetycznych i diagenetycznych (Tabela uzupełniająca 7 ). Widma nakładające się na pozycje aminokwasów z różnicami sekwencji między neandertalczykami, denisowianami, denisowianami z Xiahe i H. sapiens zostały sprawdzone ręcznie w celu ustalenia przypisania filogenetycznego. Tutaj widma z niepełną fragmentacją wokół miejsc zainteresowania zostały wyłączone z rozważań. Nie zidentyfikowano żadnych widm o wysokim poziomie ufności, co uniemożliwia przypisanie TNH2-1 do żadnej z tych trzech grup na podstawie danych paleoproteomicznych. Podobna inspekcja miejsc filogenetycznie informacyjnych wśród wielkich małp wskazuje, że TNH2-1 reprezentuje hominina (Rys. uzupełniający 14 ; Tabela uzupełniająca 8 ).
Modyfikacje białek, które zostały wcześniej zidentyfikowane w szkliwie wczesnego plejstocenu 34, zostały skwantyfikowane przez względne zliczanie widm. Dlatego liczba dopasowań peptyd-spektrum (PSM) zawierających określony PTM została podzielona przez całkowitą liczbę PSM zawierających odpowiednie aminokwasy, które można zmodyfikować. Analizę motywu sekwencji peptydów zawierających fosforylowaną resztę seryny wykonano przy użyciu pakietu R „ggseqlogo” 135 . Skrypty R do analiz PTM i konstrukcji Supplementary Fig. 4 są dostępne w danych uzupełniających 1 . Widma fragmentów peptydów obejmujące miejsca istotne dla umiejscowienia filogenetycznego zostały adnotowane przy użyciu narzędzia internetowego „Interactive Peptide Spectral Annotator” 136 (Supplementary Fig. 14 ; Supplementary Data 1 ) i skalibrowanych list pików z Maxquant. Surowe dane i identyfikacje peptydów uzyskane w wyniku naszych analiz MaxQuant są dostępne na platformie ProteomeXchange 137 (PXD018721).
Powiązana fauna z jaskini Tam Ngu Hao 2
Opis
Trzydzieści osiem zidentyfikowanych zębów (jeden mleczny i 37 stałych) suidów i liczne małe fragmenty wykazują morfologiczny wzór Sus z guzkami bunodontowymi i kłami na zębach trzonowych oraz licznymi guzkami dodatkowymi. Tylko 12 zębów (większość przedtrzonowych) z 38 jest kompletnych, przy czym większość jest fragmentaryczna lub złamana. Nie zaobserwowaliśmy znaczących różnic w zębach, które sugerowałyby obecność kilku gatunków. Na dwóch dobrze zachowanych m3 (szczególnie kompletnym TNH2-117) (ryc. 6j–k ), przednia obręcz jest zaznaczona, a środkowe kły dodatkowe są dobrze rozwinięte. Jest to również zauważalne na jednym M3 (TNH2-125), z dobrze rozwiniętą obręczą i środkowym guzkiem (hiperkonulą) za pierwszym rzędem, jak u Sus scrofa ( w porównaniu z Sus barbatus 138 ). Wymiary zmierzone na kilku zębach mieszczą się w zakresach Sus scrofa od późnego środkowego plejstocenu do późnego plejstocenu w stanowiskach indochińskich28 , 139 , 140 , 141 , w przeciwieństwie do Sus lydekkeri, który był znacznie większy25 ( Dane uzupełniające 2 ).


a – b TNH2-184, fragment prawej D4 Dicerorhinus sp. w widoku przednim ( a ) i zwarciowym ( b ). c – e TNH2-170, fragment prawej d4 (zarodek) Rhinoceros sp. w widoku policzkowym ( c ), językowym ( d ) i zwarciowym ( e ). f – g TNH2-183, fragment lewej d2 Rhinoceros sondaicus w widoku zwarciowym ( f ) i zwarciowym ( g ). h TNH2-177, fragment lewej m1/m2 Rhinoceros sp. w widoku zwarciowym. i TNH2-94, fragment lewej szczęki niezidentyfikowanej kozy w widoku policzkowym. j – k TNH2-117, prawy m3 Sus scrofa w widoku policzkowym ( j ) i zwarciowym ( k ). l TNH2-126, prawy M2 Sus scrofa w widoku zwarciowym. m – n TNH2-190, lewy p3/p4 małego jeleniowatego w widoku policzkowym ( m ) i zwarciowym ( n ). o – p TNH2-73, lewy M1/M2 kozy w widoku dystalnym ( o ) i zwarciowym ( p ). q – r TNH2-79, prawy p3 Bos sp. w widoku policzkowym ( q ) i zwarciowym ( r ). s – t TNH2-197, lewy p4 dużego jeleniowatego w widoku policzkowym ( s ) i zwarciowym ( t ). u TNH2-41, górny lewy trzonowiec jeleniowatego dużego rozmiaru w widoku zwarciowym. v – w TNH2-46, prawy d4 Bos sp. w widoku policzkowym ( v ) i zwarciowym ( w ). x – y TNH2-61, lewy p2 Bos sp. w widoku zwarciowym ( x ) i zwarciowym ( y ). z – aa TNH2-11, fragment prawego m3 Bos sp. w widoku policzkowym ( z ) i zwarciowym ( aa ). ab – ac TNH2-196, lewy d4 średniej wielkości jeleniowatego w widoku policzkowym ( ab ) i zwarciowym ( ac ). ad – aeTNH2-32, prawy D2 średniej wielkości jeleniowatego w widoku językowym ( ad ) i zwarciowym ( ae ). af – ag TNH2-167, prawy p4 Macaca sp. w widoku policzkowym ( af ) i zwarciowym ( ag ). ah TNH2-152, lewy dolny kieł samicy Macaca sp. w widoku bocznym. ai TNH2-155, lewy M1 Ursus thibetanus w widoku zwarciowym. Skala: 1 cm.
Obraz w pełnym rozmiarze
Trzydzieści pięć stałych zębów dużych bovidów jest masywnych ze stosunkowo grubym szkliwem (cecha, która pomaga odróżnić bovidy od jeleniowatych o porównywalnej wielkości, a wśród dużych bovidów, Bos od Bubalus , w oparciu o nasze własne obserwacje). Większość zębów jest fragmentaryczna, a w zestawie zębów stałych tylko trzy (jeden prawy p3, jeden lewy p2 i jeden dolny prawy m3) dostarczają danych systematyki. P2 jest mały i masywny jak u Bos , w przeciwieństwie do wydłużonego jak u Bubalus (ryc. 6x, y ). Doliny są również płytkie jak u Bos . Parastylid i parakonid p3 są mniej rozwinięte w porównaniu z kłami tylnymi. Metakonid jest zorientowany tylnie. Wymiary p2s i p3s mieszczą się w zakresach wielkości współczesnego Bos gaurus (ryc. 27 z ref. 142 ). Na m3 zauważamy wystający entostylid, który tworzy głębokie wcięcie z hypoconulidem. Nawet jeśli dane morfologiczne i morfometrycze nie są bezpośrednio porównywalne, zarys i orientacja hypoconulida na m3 są bliższe tym z późnego plejstocenu Bos gaurus frontalis (ryc. S4c z ref. 143 ) (dla tego taksonu dostępne są różne nomenklatury: Bos gaurus 144 ; Bos frontalis 145 , 146 , 147 ). Pięć fragmentarycznych zębów mlecznych i jeden kompletny lewy d3 prawdopodobnie są związane z tym zestawem zębów stałych. Duży bovid TNH2 można przypisać do Bos sp., prawdopodobnie B. cf. frontalis .
Do koziorożców z jaskini TNH2 należy również jedna mała koza z trzema prawie kompletnymi i trzema fragmentarycznymi izolowanymi zębami. Jedna fragmentaryczna szczęka nadal pokryta twardym osadem brekcyjnym pokazuje lewą stronę z D4, M1 i M2 nadal w kości (TNH2-94) (ryc. 6i ). Inny fragment zachowuje prawą stronę z M2 jako zarodkiem (TNH2-95). Cały ten materiał pokazuje archetypowy wzór podrodziny z dość gładkim szkliwem (nie tak wyszczerbionym jak u jeleniowatych) oraz szyjkami i rysikami, które są bardziej widoczne niż główne guzki i kły. Jeden dolny trzonowiec pokazuje ektostylidę, która nie jest powszechna u kóz, o ile nam wiadomo, i została zaobserwowana u jednego okazu współczesnego Naemorhedus goral z Indii (okaz MNHN 1939–243). Wymiary trzech zębów trzonowych są zgodne z zakresami rozmiarów małego gatunku Naemorhedus (ryc. 34 z piśmiennictwa 142 ), którego identyfikacja wewnątrzgatunkowa pozostaje niemożliwa na podstawie dostępnych szczątków (dane uzupełniające 2 ).
Dwadzieścia trzy zęby (19 stałych i 4 mleczne) wykazują wzór morfologiczny i wymiary podobne do zębów dużego jelenia: zryte szkliwo, duże guzki i kły na zębach trzonowych, dobrze rozwinięte ektostylidy i entostyl odpowiednio na dolnych i górnych zębach trzonowych (ryc. 6u ). Cztery dobrze zachowane dolne przedtrzonowce, jeden fragmentaryczny p2, dwa p3s i jeden p4 mogą należeć do Rusa unicolor . P4 nie jest w pełni zmolaryzowany i wykazuje częściowe zamknięcie doliny (utworzonej przez metakonid i parakonid) (ryc. 6s–t ). Ogólnie rzecz biorąc, wymiary zębów (dane uzupełniające 2 ) są małe i bardziej zbliżone do zębów dużego jeleniowatego znalezionego w Coc Muoi (późny środkowy plejstocen, Wietnam 28 ) i do zębów ze stanowiska Khok Sung (środkowy plejstocen, Tajlandia 142 ), niż do zębów ze stanowiska Tam Hay Marklot (późny plejstocen, Laos 141 ).
Zestaw rozpoznawalnych małych zębów parzystokopytnych, najbardziej fragmentarycznych, sugeruje obecność w zespole małych i średnich jeleniowatych (ryc. 6m–n, ab–ae ). Spośród dwudziestu czterech zębów, jeden m3 i jeden m1/m2, jeden wydłużony dolny p2 (parastylid prosty, metakonid zorientowany do tyłu) i dwa p3/p4, wszystkie zużyte, jeden wydłużony d2 i dwa D4, wraz z kilkoma P3/P4 mają morfologię i wymiary zgodne z Muntiacus (dane uzupełniające 2 ). Identyfikacja na poziomie gatunku jest jednak niemożliwa przy dostępnym materiale. Sześć zębów najprawdopodobniej należy do średniej wielkości jeleniowatego, wśród których dwa zużyte i fragmentaryczne zęby trzonowe, jeden siekacz, jeden d4 (TNH2-196), jeden niekompletny ząb mleczny i jeden D2 (TNH2-32). Pod względem morfologicznym i metrycznym jest on zbliżony do małego jeleniowatego z Tam Hay Marklot, wstępnie zidentyfikowanego jako Rucervus eldii (późny plejstocen, Laos 141 ), a także do Coc Muoi (późny środkowy plejstocen, Wietnam 28 ).
Próbka rhinocerotidae obejmuje głównie fragmenty izolowanych zębów. Dziewięć stałych zębów (P/p i M/m) i trzy mleczne zęby są zbyt fragmentaryczne, aby umożliwić dokładniejszą identyfikację niż Rhinoceros sp. (TNH2-170, TNH2-177 i TNH2-179) lub Rhinocerotina indet. (tj. Rhinoceros lub Dicerorhinus ) (ryc. 6c–e, h ). Niemniej jednak morfologia i wymiary dziewięciu innych okazów są bardziej pouczające. Wśród nich dwa niekompletne zęby mają cechy morfologiczne i wymiary w pełni zgodne z Rhinoceros sondaicus : i) lewy p3 (TNH2-188) z krótką, grubą i gładką wargową cingulidą oraz płytkim rowkiem ektolopidowym, idealnie pasującym do szczęki nr 305c z Soember Waroe (środkowy plejstocen, Jawa; ryc. 13 z pozycji 148 ) i p3 CM1027 z Coc Muoi (późny środkowy plejstocen, Wietnam 28 ); oraz ii) lewy d2 (TNH183), który jest wąski, z pękatym paralopidem i bez rowka przedwargowego, mieszczącego się w zakresie dużej próbki R. sondaicus z Coc Muoi 28 . Mały i fragmentaryczny prawy D4 (TNH2-184) z esowatym protolopem i zwężonym do przodu protokonem dokumentuje Dicerorhinus sp., ale jego fragmentaryczny stan wyklucza jakiekolwiek przypisanie na poziomie gatunku. Jeden tapir ( Tapirus sp.) uzupełnia różnorodność taksonomiczną perissodactyl w jaskini TNH2, z tylnym wypukłym wyrostkiem (hypolophid) fragmentarycznego d4 (TNH2-178), który jest zbyt mały, aby przypisać go do Megatapirus (szerokość: 19,5 mm zamiast 22–23 mm w ostatnim taksonie) i jest zamiast tego zgodny zarówno z T. indicus indicus , jak i T. i. intermedius 25 . Kilka fragmentów szkliwa, zarówno grubych, jak i pomarszczonych, jednoznacznie wskazuje na obecność stegodon elephantiform ( Stegodon sp.).
Małe Carnivora są reprezentowane przez dwa przedtrzonowce, jeden fragmentaryczny (TNH2-9) i jeden kompletny (TNH2-22), ale ich identyfikacja jest niemożliwa, nawet na poziomie rodzaju. Jeden dolny kieł może należeć do małego kotowatego. Dystalna część (talonid) jednego dolnego p4 (TNH2-165) jest charakterystyczna dla Viverridae, prawdopodobnie Paradoxurus sp.
Duże drapieżniki są udokumentowane za pomocą niewielu zębów, wśród których dwa kły (TNH2-159 i 182) są fragmentaryczne (Carnivora indet.). Jeden fragmentaryczny m3 i jeden zużyty M1 (TNH2-155) są przypisane do Ursus thibetanus (Fig. 6ai ). Rozmiar M1 jest zbliżony do rozmiaru Ursus thibetanus kokeni z Yenchingkuo (środkowy plejstocen, Chiny 25 ) i nieznacznie większy niż M1 z Duoi U’Oi (późny plejstocen, Wietnam 139 ) lub Coc Muoi (późny środkowy plejstocen, Wietnam 28 ). Jednak porównanie to opiera się na małych zestawach danych (mniej niż trzy okazy w każdym miejscu). Chociaż niekompletne, dwa zęby można przypisać do Ailuropoda sp. (TNH2-154 i 152). Przypisanie innego małego fragmentu do tego rodzaju jest wątpliwe (TNH2-153).
Dziesięć zębów należy do cercopithecidae ( Macaca sp.). Wymiary zmierzone na najbardziej kompletnych zębach (Dane uzupełniające 2 ), jednym p4 (TNH2-167) (Ryc. 6af–ag ), jednym M3 (TNH2-162), jednym M1/M2 (TNH2-205) i jednym m3 (TNH2-163), mieszczą się w zakresach dwóch średniej wielkości gatunków M. mulatta i M. nemestrina (Ryc. 4 z ref. 149 ) na M3 i m3; dane własne na p4). Dobrze zachowany prawy p4 (TNH2-167) przedstawia małą przednią miskę zorientowaną językowo i dużą i zaokrągloną tylną miskę jak u M. nemestrina . Ze względu na mały zestaw danych ostrożnie przypisaliśmy okazy do Macaca cf. nemestrina . Jeden fragment zęba trzonowego również świadczy o obecności orangutana ( Pongo sp.) na stanowisku Tam Ngu Hao.
Osiemnaście zębów, w większości startych, należy do dużego jeżozwierza ( Hystrix sp.) (Dane uzupełniające 2 ). Długość dwóch prawych p4, dwóch prawych m1/m2 i czterech górnych zębów trzonowych jest średnio większa niż u współczesnego azjatyckiego jeżozwierza Hystrix brachyura , a jeden m1/m2 (TNH2-143) jest nawet większy niż u innych współcześnie żyjących gatunków Hystrix indica (Ryc. 4 z ref. 150 ). Ogólnie rzecz biorąc, wymiary zębów mieszczą się w zakresie plejstoceńskiego Hystrix z kilku chińskich i indochińskich lokalizacji. Przypisanie do stosunkowo dużego gatunku H. kiangsenensis nie może zostać potwierdzone.
Komentarze dotyczące fauny towarzyszącej jaskini Tam Ngu Hao 2
Pomimo słabego zachowania odzyskanych okazów, zespół jaskiniowy Tam Ngu Hao 2 jest w dużej mierze podobny do tych znanych z południowych Chin i północnych Indochin, a w mniejszym stopniu z Jawy w środkowym plejstocenie 25 , 26 , 27 , 151 , 152. Szacowany przedział wiekowy fauny (151 +/− 37 tys. lat) oparty na łączonym datowaniu US-ESR trzech zębów w pełni zgadza się z kontekstem biochronologicznym podanym przez zespół fauny.
Podsumowanie raportu
Więcej informacji na temat projektu badawczego można znaleźć w podsumowaniu badań Nature Research Reporting Summary dostępnym pod tym artykułem.
Dostępność danych
Dane proteomiczne spektrometrii masowej wygenerowane w tym badaniu zostały złożone w ProteomeXchange Consortium za pośrednictwem repozytorium partnera PRIDE z identyfikatorem zestawu danych PXD018721 , w tym użyta baza danych referencyjnych proteomicznych. Dane pomiarowe zębów hominidów i zębów fauny wygenerowane w tym badaniu są zawarte w pliku Supplementary Data 2. Skrypt do bayesowskiego modelowania szacunków wieku Tam Ngu Hao 2 znajduje się w Supplementary Fig. 13. Ząb TNH2-1 i wszystkie zęby fauny TNH2-2 do TNH2-208 znajdują się w Muzeum Narodowym pod nadzorem Ministerstwa Informacji i Kultury Laotańskiej Republiki Ludowo-Demokratycznej. Powierzchniowy skan zęba THN2-1 i jego EDJ jest publicznie dostępny w archiwum Human Record ( https://human-fossil-record.org ), zarejestruj konto, a następnie kliknij łącze Museum of Lao PDR po lewej stronie strony). Dane źródłowe podano w niniejszym artykule.
Dostępność kodu
Kod użyty do bayesowskiego modelowania szacunków wieku Tam Ngu Hao 2 przedstawiono w postaci rysunku uzupełniającego 13 .
Odniesienia
-
Baab, KL W: Handbook of Paleoanthropology 2. (red. Henke, W. i Tattersall, I.) 2189–2219 (Springer, 2015).
-
Kaifu, Y. i in. Pokrewieństwa taksonomiczne i historia ewolucyjna wczesnoplejstoceńskich hominidów z Jawy: dowody dentognatyczne. Am. J. Phys. Anthropol. 128 , 709–726 (2005).
-
Schwartz, JH i Tattersall, I. Zapis kopalny człowieka: morfologia czaszkowo-zębowa rodzaju Homo (Afryka i Azja), 2. (Wiley-Liss, Inc., 2003).
-
Athreya, S. i Wu, X. Wielowymiarowa ocena czaszki hominina z Dali z Chin: pokrewieństwa morfologiczne i implikacje dla ewolucji plejstocenu w Azji Wschodniej. Am. J. Phys. Anthropol. 164 , 679–701 (2017).
-
Bae, CJ Późno środkowoplejstoceński zapis kopalny homininów w Azji Wschodniej: synteza i przegląd. Am. J. Phys. Anthropol. 143 , 75–93 (2010).
-
Xing, S. i in. Zęby homininów z wczesnego, późnoplejstoceńskiego stanowiska Xujiayao w północnych Chinach. Am. J. Phys. Anthropol. 156 , 224–240 (2015).
-
Roksandic, M., Radović, P., Wu, X.-J. & Bae, CJ Rozwiązywanie „zamieszania pośrodku”: przypadek Homobodoensis sp. nov. Evol. Anthropol . (2021). https://doi.org/10.1002/evan.21929 .
-
Ji, Q., Wu, W., Ji, Y., Li, Q. i Ni, X. Czaszka Harbin z późnego środkowego plejstocenu reprezentuje nowy. Homo Species Innov. 2 , 100132 (2021).
-
Li, Z.-Y. i in. Późnoplejstoceńskie archaiczne ludzkie czaszki z Xuchang w Chinach. Science 355 , 969–972 (2017).
-
Wu, XJ i in. Archaiczne szczątki ludzkie z Hualongdong w Chinach oraz ciągłość i zmienność człowieka w środkowym plejstocenie. Proc. Natl Acad. Sci. USA 116 , 9820–9824 (2019).
-
Stringer, C. Status Homo heidelbergensis (Schoetensack 1908). Evol. Anthropol. 21 , 101–107 (2012).
-
Liu, W., Wu, XJ, Xing, S. i Zhang, YY Skamieniałości ludzkie w Chinach (Science Press, 2014).
-
Krause, J. i in. Kompletny genom DNA mitochondrialnego nieznanego hominina z południowej Syberii. Nature 464 , 894–897 (2010).
-
Sawyer, S. i in. Sekwencje DNA jądrowego i mitochondrialnego dwóch osobników z plemienia Denisova. Proc. Natl Acad. Sci. USA 112 , 15696–15700 (2015).
-
Chen, F. i in. Późno środkowoplejstoceńska żuchwa denisowiana z Wyżyny Tybetańskiej. Nature 569 , 409–412 (2019).
-
Slon, V. i in. Czwarty osobnik Denisovana. Nauka. Adw. 3 , e1700186 (2017).
-
Jacobs, Z. i in. Czas okupacji archaicznej homininy w Jaskini Denisowej w południowej Syberii. Natura 565 , 594–599 (2019).
-
Teixeira, JC i in. Powszechne pochodzenie denisowiańskie na wyspach Azji Południowo-Wschodniej, ale brak dowodów na znaczną domieszkę superarchaicznych homininów. Nat. Ecol. Evol. 5 , 616–624 (2021).
-
Larena, M. i in. Filipińska Ayta posiada najwyższy poziom pochodzenia denisowskiego na świecie. Curr. Biol. 31 , 4219–4230 (2021).
-
Demeter, F. i in. Anatomicznie współczesny człowiek w Azji Południowo-Wschodniej (Laos) do 46 r. n.e. Proc. Natl Acad. Sci. USA 109 , 14375–14380 (2012).
-
Demeter, F. i in. Wcześni ludzie współcześni i zmienność morfologiczna w Azji Południowo-Wschodniej: dowody kopalne z Tam Pa Ling w Laosie. PLoS One 10 , e0121193 (2015).
-
Shackelford, L. i in. Dodatkowe dowody na wczesną nowożytną różnorodność morfologiczną człowieka w Azji Południowo-Wschodniej w Tam Pa Ling, Laos. Quat. Int. 466 , 93–106 (2018).
-
Al Qahtani, SJ i Liversidge, HM Londyński atlas rozwoju i wyrzynania się zębów ludzkich. Am. J. Phys. Anthropol. 142 , 481–490 (2010).
-
Bacon, AM i in. Zespoły ssaków z późnego plejstocenu w Azji Południowo-Wschodniej: nowe datowanie, profile śmiertelności i ewolucja relacji drapieżnik-ofiara w kontekście środowiskowym. Palaeogeogr. Palaeoclimatol. Palaeoecol. 422 , 101–127 (2015).
-
Colbert, EH i Hooijer, DA Ssaki plejstoceńskie z wapiennych szczelin Syczuanu w Chinach. Bull. Amer. Mus. Nat. Hist. 102 , 1–134 (1953).
-
Wang, W. i in. Sekwencja skamieniałości ssaków, w tym zęby hominoidów, z jaskiń basenu Bubing w południowych Chinach. J. Hum. Evol. 52 , 370–379 (2007).
-
Rink, WJ, Wei, W., Bekken, D. & Jones, HL Geochronologia fauny Ailuropoda-Stegodon i Gigantopithecus w prowincji Guangxi w południowych Chinach. Kwat. Rozdzielczość 69 , 377–387 (2008).
-
Bacon, A.-M. i in. Ponowne odwiedzenie stanowisk w Azji Południowo-Wschodniej w Nam Lot (MIS 5) i Duoi U’Oi (MIS 4): dowody zooarcheologiczne i izotopowe. Palaeogeogr. Palaeoclimatol. Palaeoecol. 512 , 132–144 (2018).
-
Ciochon, R. i in. Datowane współwystępowanie Homo erectus i Gigantopithecus z jaskini Tham Khuyen w Wietnamie. Proc. Natl Acad. Sci. USA 93 , 3016–3020 (1996).
-
Zhang, Y. i in. Nowe szczątki Gigantopithecus blacki z 400–320 tys. lat z jaskini Hejiang, miasto Chongzuo, Guangxi, południowe Chiny. Quat. Int. 342 , 35–45 (2014).
-
Bocherens, H. i in. Elastyczność diety i siedliska u plejstoceńskich ssaków południowoazjatyckich: Implikacje dla losu gigantycznego małpiatka kopalnego Gigantopithecus. Quat. Int. 434A , 148–155 (2017).
-
Turvey, ST, Tong, H., Stuart, AJ i Lister, AM Holocene survival of Late Pleistocene megafauna in China: a critical review of the evidence. Quat. Sci. Rev. 76 , 156–166 (2013).
-
Bacon, AM i in. Wieloproxyjność podejścia do eksploracji przybycia, środowisk i adaptacji Homo sapiens w Azji Południowo-Wschodniej. Sci. Rep. 11 , 1–14 (2021).
-
Cappellini, E. i in. Wczesnoplejstoceński proteom szkliwa z Dmanisi rozstrzyga filogenezę Stephanorhinus . Nature 574 , 103–107 (2019).
-
Welker, F. i in. Proteom zębów Homo antecessor . Nature 580 , 235–238 (2020).
-
Welker, F. i in. Proteom szkliwa pokazuje, że Gigantopithecus był wczesnym rozbieżnym pongine. Nature 576 , 262–265 (2019).
-
Bailey, SE i Hublin, J.-J. W: Anthropological Perspectives on Tooth Morphology (red. Scott, GR i Irish, JD) 222–249 (Cambridge University Press, 2013).
-
Bailey, SE, Skinner, MM i Hublin, JJ Co kryje się pod spodem? Ocena wzorów grzebienia trygonidów dolnych zębów trzonowych na podstawie ekspresji zębiny i szkliwa. Am. J. Phys. Anthropol. 145 , 505–518 (2011).
-
Skinner, MM i in. Stomatologiczna perspektywa powinowactwa taksonomicznego żuchwy Balanica (BH-1). J. Hum. Evol. 93 , 63–81 (2016).
-
Olejniczak, AJ i in. Morfologia połączenia szkliwa z zębiną w przekrojach zębów trzonowych szczęki naczelnych człekokształtnych. J. Hum. Evol. 53 , 292–301 (2007).
-
Zanolli, C. Organizacja wewnętrznej struktury korony zęba trzonowego u Homo erectus jawajskiego . Am. J. Phys. Anthropol. 156 , 148–157 (2015).
-
Zanolli, C. i in. Dowody na zwiększoną różnorodność hominidów we wczesnym i środkowym plejstocenie w Indonezji. Nat. Ecol. Evol. 3 , 755–764 (2019).
-
Zanolli, C. i in. Szczątki zębów człowieka z późnego wczesnego plejstocenu z Uadi Aalad i Mulhuli-Amo (Buia), erytrejski Danakil: makromorfologia i mikrostruktura. J. Hum. Evol. 74 , 96–113 (2014).
-
Zanolli, C. i Mazurier, A. Charakterystyka endostrukturalna szczątków zębowych H. heidelbergensis z wczesnego środkowoplejstoceńskiego stanowiska Tighenif w Algierii. Comptes Rendus Palevol 12 , 293–304 (2013).
-
Xing, S., Martinón-Torres, M. i Bermúdez De Castro, JM Skamieniałe zęby człowieka pekińskiego. Nauka. Rep. 8 , 2066 (2018).
-
Xing, S. i in. Zęby homininów ze stanowiska środkowoplejstoceńskiego w Yiyuan we wschodnich Chinach. J. Hum. Evol. 95 , 33–54 (2016).
-
Xing, S. i in. Zęby homininów ze środkowego plejstocenu z jaskini Longtan, Hexian, Chiny. PLoS One 9 , 1–38 (2014).
-
Pan, L., Dumoncel, J., Mazurier, A. i Zanolli, C. Różnorodność homininów w Azji Wschodniej w środkowym plejstocenie: perspektywa endostrukturalna przedtrzonowców. J. Hum. Evol. 148 , 102888 (2020).
-
Morwood, MJ i in. Dalsze dowody na istnienie homininów o małych ciałach z późnego plejstocenu Flores w Indonezji. Natura 437 , 1012–1017 (2005).
-
Détroit, F. i in. Nowy gatunek Homo z późnego plejstocenu Filipin. Nature 568 , 181–186 (2019).
-
Jacobs, GS i in. Wielorakie głęboko rozbieżne przodkowie denisowiańscy u Papuasów. Komórka 177 , 1010–1021.e32 (2019).
-
Slon, V. i in. Genom potomstwa matki neandertalczyka i ojca denisowianina. Nature 561 , 113–116 (2018).
-
Browning, SR, Browning, BL, Zhou, Y., Tucci, S. i Akey, JM Analiza danych sekwencji ludzkich ujawnia dwa impulsy archaicznej domieszki denisovańskiej. Komórka 173 , 53–61.e9 (2018).
-
Gokhman, D. i in. Rekonstrukcja anatomii denisovana przy użyciu map metylacji DNA. Cell 179 , 180–192.e10 (2019).
-
Chang, CH i in. Pierwszy archaiczny Homo z Tajwanu. Nat. Commun. 6 , 1–33 (2015).
-
Wu, X. & Poirier, FE Ewolucja człowieka w Chinach. Metryczny opis skamieniałości i przegląd stanowisk . (Oxford University Press, 1995).
-
Xiao, D. i in. Metryczna i geometryczna analiza morfometryczna nowych skamieniałości homininów z Maba (Guangdong, Chiny). J. Hum. Evol. 74 , 1–20 (2014).
-
Rizal, Y. i in. Ostatnie pojawienie się Homo erectus w Ngandong na Jawie 117 000–108 000 lat temu. Natura 577 , 381–385 (2020).
-
Morley, MW i in. Wstępne wyniki mikromorfologiczne z Liang Bua, Flores (Indonezja): Procesy formowania się miejsc i aktywność homininów w lokalizacji typowej Homo floresiensis . J. Archaeol. Sci. 77 , 125–142 (2017).
-
Morley, MW i in. Aktywności hominidów i zwierząt w zapisie mikrostratygraficznym z jaskini Denisova (góry Ałtaj, Rosja). Sci. Rep. 9 , 13785 (2019).
-
Stoops, G. Wytyczne dotyczące analizy i opisu cienkich przekrojów gleby i regolitu . (Soil Science Society of America, 2003).
-
McAdams, C. i in. Plejstoceńska geoarcheologia i geochronologia jaskini Con Moong w północnym Wietnamie: procesy formowania się stanowisk i aktywność homininów w wilgotnych tropikach. Geoarcheologia 35 , 72–97 (2019).
-
Herries, AIR i in. Współczesność Australopithecus , Paranthropus i wczesnego Homo erectus w Afryce Południowej. Science 368 , eaaw7293 (2020).
-
Kolobova, KA i in. Archeologiczne dowody na dwa oddzielne rozproszenia neandertalczyków w południowej Syberii. Proc. Natl Acad. Sci. USA 117 , 2879–2885 (2020).
-
Aitken, MJ Wprowadzenie do datowania optycznego: datowanie osadów czwartorzędowych metodą luminescencji stymulowanej fotonami (Oxford University Press, 1998).
-
Hutt, G., Jaek, I. i Tchonka, J. Datowanie optyczne: widma stymulacji odpowiedzi optycznej skalenia potasowego. Quat. Sci. Rev. 7 , 381–385 (1988).
-
Thomsen, KJ, Murray, AS, Jain, M. i Botter-Jensen, L. Laboratoryjne tempo zanikania różnych sygnałów luminescencji z ekstraktów osadów bogatych w skalenie. Radiat. Meas. 43 , 1474–1486 (2008).
-
Bøtter-Jensen, L. & Mejdahl, V. Ocena mocy dawki beta przy użyciu wielopunktowego systemu GM. Int. J. Radiat. Appl. Instrum. Część 14 , 187–191 (1988).
-
Brennan, BJ Dawki beta do ziaren kulistych. Radiat. Meas. 37 , 299–303 (2003).
-
Wang, W. i Xia, J. Pomiar rocznej dawki z łańcuchów rozpadu serii Th i U metodą zliczania fal alfa z grubego źródła. Archaeometry1 14 , 101–108 (1991).
-
Guerin, G., Mercier, N. i Adamiec, G. Współczynniki przeliczeniowe mocy dawki: aktualizacja. Anc. TL 29 , 5–11 (2011).
-
Huntley, DJ i Baril, MR Zawartość potasu w skaleniach potasowych mierzona metodą datowania optycznego lub termoluminescencyjnego. Anc. TL 15 , 11–13 (1997).
-
Huntley, DJ i Hancock, RGV Zawartość Rb w skaleniach potasowych mierzonych w datowaniu optycznym. Anc. TL 19 , 43–46 (2001).
-
Prescott, JR i Hutton, JT Wkład promieniowania kosmicznego w moce dawek dla luminescencji i datowania ESR: duże głębokości i długoterminowe zmiany w czasie. Radiat. Meas. 23 , 497–500 (1994).
-
Stokes, S. i in. Alternatywne chronologie dla późnoczwartorzędowych (ostatni interglacjał-holocen) głębokomorskich osadów poprzez datowanie optyczne kwarcu wielkości mułu. Quat. Sci. Rev. 22 , 925–941 (2003).
-
Roberts, HM Testowanie protokołów IRSL po IR w celu zminimalizowania blaknięcia skaleni, przy użyciu lessu alaskańskiego z niezależną kontrolą chronologiczną. Radiat. Meas. 47 , 716–724 (2012).
-
Rhodes, EJ Datowanie osadów przy użyciu skalenia potasowego jednoziarnistego IRSL: wstępne rozważania metodologiczne. Quat. Int. 362 , 14–22 (2015).
-
Buylaert, JP, Murray, AS, Thomsen, KJ i Jain, M. Testowanie potencjału sygnału IRSL o podwyższonej temperaturze z potasowego skalenia. Radiat. Meas. 44 , 560–565 (2009).
-
Murray, AS, Buylaert, JP, Thomsen, KJ i Jain, M. Wpływ wstępnego podgrzewania na sygnał IRSL ze skalenia. Radiat. Meas. 44 , 554–559 (2009).
-
Thiel, C. i in. Datowanie luminescencyjne profilu lessowego Stratzing (Austria) — testowanie potencjału protokołu post-IR IRSL w podwyższonej temperaturze. Quat. Int. 234 , 23–31 (2011).
-
Thomsen, KJ, Murray, AS i Jain, M. Stabilność sygnałów IRSL z próbek osadowych skalenia potasowego. Geochronometria 38 , 1–13 (2011).
-
Li, B. i Li, S. Odpowiedź na komentarze Thomsena i in. na temat „Datowanie luminescencyjne potasowego skalenia z osadów: protokół bez korekty zanikania anomalii”. Quat. Geochronol. 8 , 49–51 (2012).
-
Brown, ND, Rhodes, EJ, Antinao, JL i McDonald, EV Sygnały pojedynczego ziarna post-IR IRSL skaleni potasowych z osadów stożków napływowych w Baja California Sur w Meksyku. Quat. Int. 362 , 132–138 (2015).
-
Huntley, DJ i Lamothe, M. Wszechobecność anomalii zanikania w skaleniach potasowych oraz jej pomiar i korekta w datowaniu optycznym. Can. J. Earth Sci. 28 , 1093–1106 (2001).
-
Jacobs, Z., Duller, GAT i Wintle, AG Interpretacja rozkładów De pojedynczego ziarna i obliczanie De. Radiat. Meas. 41 , 264–277 (2006).
-
Olley, JM, Murray, A. i Roberts, RG Wpływ nierównowagi w łańcuchach rozpadu uranu i toru na moce dawek zakopanych w osadach rzecznych. Quat. Sci. Rev. 15 , 751–760 (1996).
-
Zhou, H., Zhao, J., Qing, W., Feng, Y. i Tang, J. Zróżnicowanie letniego monsunu azjatyckiego w środkowych Chinach spowodowane speleothemami. 54–46 tys. lat temu. J. Quat. Sci. 26 , 781–790 (2011).
-
Clark, TR i in. Rozróżnianie czasu i przyczyny historycznych zdarzeń śmiertelności w nowoczesnych pority z Wielkiej Rafy Koralowej. Geochim. Cosmochim. Acta 138 , 57–80 (2014).
-
Ludwig, KR Podręcznik użytkownika programu Isoplot 3.75. Zestaw narzędzi geochronologicznych dla programu Microsoft Excel (Berkeley Geochronology Center Special Publication No. 5, 2012).
-
Cheng, H. i in. Okresy półtrwania uranu-234 i toru-230. Chem. Geol. 169 , 17–33 (2000).
-
Eggins, SM i in. Datowanie in situ U-series metodą ablacji laserowej wielokolektorowego ICPMS: nowe perspektywy dla geochronologii czwartorzędu. Quat. Sci. Rev. 24 , 2523–2538 (2005).
-
Joannes-Boyau, R. & Grün, R. Zachowanie termiczne zorientowanych i nie zorientowanych rodników CO2- w szkliwie zębów. Radiat. Meas. 44 , 505–511 (2009).
-
Joannes-Boyau, R., Grün, R. i Bodin, T. Rozkład składowej promieniowania gamma w laboratorium w widmach kątowych ESR fragmentów szkliwa zębów kopalnych. Appl. Radiat. Isot. 68 , 1798–1808 (2010).
-
Joannes-Boyau, R. Szczegółowy protokół dokładnego nieniszczącego datowania bezpośredniego fragmentu szkliwa zęba przy użyciu rezonansu spinowego elektronów. Geochronometria 40 , 322–333 (2013).
-
Grün, R., Athreya, S., Raj, R. i Patnaik, R. Odpowiedź ESR w szkliwie zębów na tomografię komputerową o wysokiej rozdzielczości. Archaeol. Anthropol. Sci. 4 , 25–28 (2012).
-
Westaway, KE i in. Wczesna obecność człowieka współczesnego na Sumatrze 73 000–63 000 lat temu. Nature 548 , 322–325 (2017).
-
Joannes-Boyau, R., Duval, M. i Bodin, T. McDoseE 2.0 Nowy program Monte Carlo oparty na łańcuchu Markowa do dopasowywania krzywej zależności dawka-odpowiedź ESR i oceny dawki. Quat. Geochronol. 44 , 13–22 (2018).
-
Joannes-Boyau, R. & Grün, R. Kompleksowy model rodników CO2- w szkliwie zębów kopalnych: Implikacje dla datowania ESR. Quat. Geochronol. 6 , 82–97 (2011).
-
Duval, M. & Grün, R. Czy opublikowane oceny dawek ESR w przypadku szkliwa zębów kopalnych są wiarygodne? Quat. Geochronol. 31 , 19–27 (2016).
-
Shao, Q. i in. ESR, seria U i datowanie paleomagnetyczne fauny Gigantopithecus z jaskini Chuifeng w Guangxi w południowych Chinach. Kwat. Rozdzielczość 82 , 270–280 (2014).
-
Bronk Ramsey, C. Kalibracja radiowęglowa i analiza stratygrafii: program OxCal. Radiocarbon 37 , 425–430 (1995).
-
Smith, BH Wzory zużycia zębów trzonowych u łowców-zbieraczy i rolników. Am. J. Phys. Anthropol. 63 , 39–56 (1984).
-
Martínez de Pinillos, M. i in. Ekspresja grzebienia trygonidów w dolnych zębach trzonowych Atapuerca-Sima de los Huesos: wewnętrzna i zewnętrzna ekspresja morfologiczna oraz wnioski ewolucyjne. Comptes Rendus Palevol 13 , 205–221 (2014).
-
Turner II, CG, Nichol, CR i Scott, GR w Advances in Dental Anthropology (red. Kelley, MA i Larsen, CS) 13–31 (Wiley-Liss, Inc., 1991).
-
Scott, GR i Irish, JD Human Tooth Crown and Root Morphology. Arizona State University Dental Anthropology System (Cambridge University Press, 2017).
-
Spoor, CF, Zonneveld, FW i Macho, GA Liniowe pomiary kości korowej i szkliwa zębowego za pomocą tomografii komputerowej: zastosowania i problemy. Am. J. Phys. Anthropol. 91 , 469–484 (1993).
-
Fajardo, RJ, Ryan, TM i Kappelman, J. Ocena dokładności tomografii komputerowej rentgenowskiej o wysokiej rozdzielczości kości beleczkowej naczelnych poprzez porównania z przekrojami histologicznymi. Am. J. Phys. Anthropol. 118 , 1–10 (2002).
-
Coleman, MN i Colbert, MW Uwaga techniczna: Protokoły progowania CT do wykonywania pomiarów na modelach trójwymiarowych. Am. J. Phys. Anthropol. 133 , 723–725 (2007).
-
Maureille, B., Rougier, H., Houët, F. & Vandermeersch, B. Les dents inférieures du néandertalien Regourdou 1 (site de Regourdou, gmina Montignac, Dordogne): analizy métriques et porównawcze. Rev. d’archéologie pr.éhistorique 13 , 183–200 (2001).
-
Scolan, H., Santos, F., Tillier, AM, Maureille, B. & Quintard, A. Des nouveaux pozostałości néanderthaliens à Las Pélénos (Monsempron-Libos, Lot-et-Garonne, Francja). Byk. Pam. Towarzystwo Antropol. Paryż 24 , 69–95 (2012).
-
Olejniczak, AJ i in. Proporcje tkanek zębowych i grubość szkliwa zębów trzonowych neandertalczyków i ludzi współczesnych. J. Hum. Evol. 55 , 12–23 (2008).
-
Olejniczak, AJ i in. Grubość szkliwa zębów trzonowych i wysokość rogu zębiny u Gigantopithecus blacki . Am. J. Phys. Anthropol. 135 , 85–91 (2008).
-
Macchiarelli, R., Bayle, P., Bondioli, L., Mazurier, A. i Zanolli, C. W: Antropologiczne perspektywy morfologii zębów: genetyka, ewolucja, zmienność (red. Scott, RG i Irish, JD) 250–277 (Cambridge University Press, 2013).
-
Durrleman, S. i in. W Medical Image Computing and Computer-Assisted Intervention – MICCAI 2012 (red. Ayache, N., Delingette, H., Golland, P. i Mori, K.) 223–230 (Springer, 2012).
-
Durrleman, S., Pennec, X., Trouvé, A., Ayache, N. i Braga, J. Porównanie ontogenezy endokranialnej szympansów i bonobo za pomocą regresji czasowej i rejestracji czasoprzestrzennej. J. Hum. Evol. 62 , 74–88 (2012).
-
Durrleman, S. i in. Morfometria kompleksów kształtu anatomicznego o gęstych deformacjach i rzadkich parametrach. Neuroimage 101 , 35–49 (2014).
-
Zanolli, C. i in. Morfologia zębów wewnętrznych Homo erectus z Zhoukoudian. Nowe dowody ze starej kolekcji przechowywanej na Uniwersytecie w Uppsali w Szwecji. J. Hum. Evol. 116 , 1–13 (2018).
-
Braga, J. i in. Skuteczność dopasowania powierzchni dyfeomorficznej i morfometrii geometrycznej 3D w dyskryminacji taksonomicznej zębów trzonowych żuchwy u homininów z wczesnego plejstocenu. J. Hum. Evol. 130 , 21–35 (2019).
-
Bookstein, FL Metody wyznaczania punktów orientacyjnych dla form bez punktów orientacyjnych: morfometria różnic grupowych w kształcie obrysu. Med. Image Anal. 1 , 225–243 (1997).
-
Gunz, P. & Mitteroecker, P. Semilandmarks: metoda kwantyfikacji krzywych i powierzchni. Hystrix 24 , 103–109 (2013).
-
Gunz, P., Mitteroecker, P. i Bookstein, FL w Modern Morphometrics in Physical Anthropology (red. Slice, D.) 73–98 (Kluwer Academic/Plenum Publishers, 2005).
-
Skinner, MM, Gunz, P., Wood, BA i Hublin, JJ Morfologia złącza szkliwo-zębina (EDJ) odróżnia dolne zęby trzonowe Australopithecus africanus i Paranthropus solidus . J. Hum. Ewolucja 55 , 979–988 (2008).
-
Skinner, MM, Gunz, P., Wood, BA, Boesch, C. i Hublin, JJ Rozróżnianie istniejących gatunków i podgatunków Pan na podstawie morfologii połączenia szkliwa z zębiną dolnych zębów trzonowych. Am. J. Phys. Anthropol. 140 , 234–243 (2009).
-
Mitteroecker, P. & Bookstein, F. Dyskryminacja liniowa, ordynacja i wizualizacja gradientów selekcji we współczesnej morfometrii. Evol. Biol. 38 , 100–114 (2011).
-
Cardini, A. i Polly, PD Krzyżowa walidacja między wykresami punktowymi PCA grup: rozwiązanie problemu błędnego rozdzielenia grup? Evol. Biol. 47 , 85–95 (2020).
-
Cardini, A., O’Higgins, P. i Rohlf, FJ Dostrzeganie odrębnych grup tam, gdzie ich nie ma: fałszywe wzorce z analizy składniowej komórek międzygrupowych. Evol. Biol. 46 , 303–316 (2019).
-
Dray, S. & Dufour, AB Pakiet ade4: implementacja diagramu dualności dla ekologów. J. Stat. Softw. 22 , 1–20 (2007).
-
Schlager, S. W: Statistical Shape and Deformation Analysis (red. Zheng, G., Li, S. i Szekely, G.) 217–256 (Academic Press, 2017).
-
Zespół, RDC R: język i środowisko do obliczeń statystycznych. http://www.r-project.org (2020).
-
Bookstein, FL Narzędzia morfometrycze do danych o obiektach orientacyjnych: geometria i biologia (Cambridge University Press, 1991).
-
Rappsilber, J., Ishihama, Y. i Mann, M. Końcówki ekstrakcyjne Stop And Go do desorpcji/jonizacji laserowej wspomaganej matrycą, nanoelektrosprayu i wstępnej obróbki próbek LC/MS w proteomice. Anal. Chem. 75 , 663–670 (2003).
-
Kelstrup, CD i in. Ocena wydajności Q-exclusive HF-X w proteomice strzelbowej. J. Proteome Res. 17 , 727–738 (2018).
-
Tyanova, S., Temu, T. i Cox, J. Platforma obliczeniowa MaxQuant do proteomiki shotgun opartej na spektrometrii masowej. Nat. Protoc. 11 , 2301–2319 (2016).
-
Mackie, M. i in. Profilowanie paleoproteomiczne warstw konserwatorskich na włoskim malowidle ściennym z XIV wieku. Angew. Chem. Int. Ed. 57 , 7369–7374 (2018).
-
Wagih, O. Ggseqlogo: wszechstronny pakiet R do rysowania logów sekwencyjnych. Bioinformatics 33 , 3645–3647 (2017).
-
Brademan, DR, Riley, NM, Kwiecien, NW i Coon, JJ Interaktywny adnotator widma peptydowego: wszechstronne narzędzie internetowe do zastosowań proteomicznych. Mol. Cell. Proteom. 18 , S193–S201 (2019).
-
Vizcaíno, JA i in. ProteomeXchange zapewnia globalnie skoordynowane przesyłanie i rozpowszechnianie danych proteomicznych. Nat. Biotechnol. 32 , 223–226 (2014).
-
Badoux, DM Ssaki kopalne z dwóch złóż w Punung (Jawa) (Uniwersytet w Utrechcie, 1959).
-
Bacon, AM i in. Redécouverte d’une faune pléistocène dans les remplissages karstiques de Tam Hang au Laos: premiery résultats. Comptes Rendus Palevol. 7 , 277–288 (2008).
-
Bacon, A.-M. i in. Środkowoplestoceńska fauna ssaków z krasowego złoża Tam Hang, północny Laos: nowe dane i hipoteza ewolucyjna. Quat. Int. 245 , 315–332 (2011).
-
Bourgon, N. i in. Izotopy cynku w późnoplejstoceńskich zębach kopalnych z jaskini w Azji Południowo-Wschodniej zachowują informacje paleodietarne. Proc. Natl Acad. Sci. USA 9 , 4675–4681 (2020).
-
Suraprasit, K. i in. Środkowopleistoceńska fauna kręgowców z Khok Sung (Nakhon Ratchasima, Tajlandia): implikacje biochronologiczne i paleobiogeograficzne. Zookeys 613 , 1–157 (2016).
-
Cucchi, T. i in. Sygnały taksonomiczne i filogenetyczne w zębach policzkowych bydła: w kierunku nowych markerów biosystemowych do eksploracji historii bydła dzikiego i udomowionego. J. Archaeol. Sci. 109 , 104993 (2019).
-
Gentry, A., Clutton-Brock, J. i Groves, CP Nazewnictwo gatunków zwierząt dzikich i ich udomowionych odpowiedników. J. Archaeol. Sci. 31 , 645–651 (2004).
-
Wilson, DL i Reeder, DM Mammal Species of the World, a Taxonomic and Geographic Reference, wydanie 3. (Johns Hopkins University Press, 2005).
-
Tougard, C. Les faunes de grands mammiferes du pleistocene Moyen Terminal de Thaie Dans Leur Cadre Pnylogenetique, Paleoecologique et Biochronologique. (Uniwersytet w Montpellier, 1998).
-
Filoux, A., Wattanapituksakul, A., Lespes, C. i Thongcharoenchaikit, C. Plejstoceńskie zgromadzenie ssaków zawierające Ailuropoda i Pongo z jaskini Tham Prakai Phet w prowincji Chaiyaphum w Tajlandii. Geobios 48 , 341–349 (2015).
-
Hooijer, DA Prehistoryczne i kopalne nosorożce z Archipelagu Malajskiego i Indii. Zool. Meded. 26 , 1–138 (1946).
-
Takai, M., Zhang, Y., Kono, RT i Jin, C. Zmiany w składzie plejstoceńskiej fauny naczelnych w południowych Chinach. Quat. Int. 354 , 75–85 (2014).
-
van Weers, DJ Rewizja taksonomiczna plejstoceńskiego Hystrix (Hystricidae, Rodentia) z Eurazji z notatkami na temat ewolucji rodziny. Contrib. Zool. 74 , 301–312 (2005).
-
Norton, CJ, Jin, C., Wang, Y. i Zhang, Y. In Vertebrate Paleobiology and Paleoanthropology (Norton, CJ i Braun, red. DR) 81–100 (Springer, 2011).
-
Bacon, AM i in. Megafauna zdominowana przez nosorożce w okresie przejściowym MIS6-5: zespół Coc Muoi z późnego środkowoplejstocenu, prowincja Lang Son, Wietnam. Quat. Sci. Rev. 186 , 123–141 (2018).
Komunikacja przyrodnicza tom 13 , Numer artykułu: 2557 ( 2022 )
Podziękowanie
Zespół dedykuje tę pracę panu Thongsa Sayavongkhamdy. Dziękujemy Ministerstwu Informacji, Kultury i Turystyki Laotańskiej Republiki Ludowo-Demokratycznej za zachęcanie i wspieranie naszej pracy, która trwa już prawie 20 lat. Dziękujemy władzom dystryktu Xon, prowincji Hua Pan i mieszkańcom wioski Long Gua Pa za ciągłe wsparcie naszych wieloletnich prac terenowych. Dziękujemy również wielu kuratorom i współpracownikom, którzy udostępnili nam porównawcze skamieniałości i niedawne okazy homininów do skanowania, a także internetowym platformom udostępniania Nespos Society ( http://www.nespos.org ) i ESRF Paleontological Database ( http://paleo.esrf.eu ). Dziękujemy A. Mazurier i R. Macchiarelli (University of Poitiers), A. Bravin, C. Nemoz i P. Tafforeau (ESRF Synchrotron), P. Bayle i F. Santos (University of Bordeaux), O. Kullmer i F. Schrenk (Senckenberg Museum), F. Bernardini i C. Tuniz (ICTP Triest), J. Braga i J. Dumoncel (University of Tuluza) za wsparcie analityczne, skanowanie mikrotomograficzne i udostępnianie materiałów. Z wdzięcznością przyjmujemy wsparcie ze strony CNRS/IN2P3 Computing Center (Lyon – Francja) za udostępnienie zasobów obliczeniowych i przetwarzania danych potrzebnych do tej pracy. Podziękowania kierujemy również do Christine Lefèvre, Joséphine Lesur i Aurélie Verguin (Laboratoire Mammifères et Oiseaux, Anatomie comparée, MNHN, Paryż) za dostęp do porównawczych kolekcji ssaków. Serdeczne podziękowania dla Jeremy’ego Kazana (Taxidermie, MNHN, Paryż) za, jak zwykle, wspaniałe odlanie i odtworzenie okazu. MWM dziękuje George’owi Morganowi i Markowi Keene’owi z Adelaide Petrographics za fachowe wykonanie cienkich przekrojów mikromorfologicznych. Badania te zostały sfinansowane przez stypendium Future Fellowship przyznane przez Australian Research Council (ARC) (FT180100309) (MWM), grant Discovery przyznany przez Australian Research Council (ARC) (DP170101597) (KEW) oraz grant LEF przyznany przez Australian Research Council (ARC) (LE200100022) (RJ-B.), stypendium Marie Skłodowska Curie Individual Fellowship (nr 795569) (FW). Marie Skłodowska-Curie European Training Network (ETN) TEMPERA, projekt finansowany przez Program Ramowy Unii Europejskiej w zakresie badań naukowych i innowacji Horyzont 2020 (nr 722606) (KE, JVO, PR), Europejską Radę ds. Badań Naukowych (ERBN) w ramach programu badań naukowych i innowacji Horyzont 2020 Unii Europejskiej (nr 948365) (FW), VILLUM FONDEN (nr 17649) (KE). Program Ramowy Unii Europejskiej w zakresie badań naukowych i innowacji Horyzont 2020 (nr 819960) (MMS), Novo Nordisk Foundation (numer grantu NNF14CC0001) (PLR), National Geographic Society (NGS-399R-18) (LS), MNHN/Université Paris Diderot/Sorbonne Paris Cité, Francja (FD), CNRS/Université de Paris, Francja; Uniwersytet w Strasburgu, Francja (AMB). Katedra Ewolucji Człowieka, Instytut Antropologii Ewolucyjnej im. Maxa Plancka w Lipsku, Niemcy (AMB) i firma Sodipram (La Compagnie du Lit), Francja (FD).
Deklaracje etyczne
Konflikty interesów
Autorzy deklarują brak konfliktu interesów.
Recenzja ekspercka
Informacje o recenzji eksperckiej
Nature Communications dziękuje Bence Viola i innym anonimowym recenzentom za ich wkład w recenzję tej pracy. Raporty recenzentów są dostępne.
Informacje dodatkowe
Uwaga wydawcy: Springer Nature zachowuje neutralność w kwestii roszczeń jurysdykcyjnych zawartych w publikowanych mapach i powiązań instytucjonalnych.
Dane źródłowe
Prawa i uprawnienia
Otwarty dostęp Niniejszy artykuł jest licencjonowany na podstawie licencji Creative Commons Uznanie autorstwa 4.0 Międzynarodowe, która zezwala na używanie, udostępnianie, adaptację, dystrybucję i reprodukcję w dowolnym medium lub formacie, pod warunkiem podania odpowiedniego uznania dla oryginalnych autorów i źródła, podania linku do licencji Creative Commons i wskazania, czy wprowadzono zmiany. Obrazy lub inne materiały stron trzecich w tym artykule są objęte licencją Creative Commons artykułu, chyba że wskazano inaczej w linii kredytowej do materiału. Jeśli materiał nie jest objęty licencją Creative Commons artykułu, a zamierzone użycie nie jest dozwolone przez przepisy ustawowe lub przekracza dozwolone użycie, należy uzyskać zgodę bezpośrednio od właściciela praw autorskich. Aby wyświetlić kopię tej licencji, odwiedź stronę http://creativecommons.org/licenses/by/4.0/ .
O tym artykule


Cytuj ten artykuł
Demeter, F., Zanolli, C., Westaway, KE i in . Środkowy plejstoceński denisowiański trzonowiec z łańcucha Annamitów w północnym Laosie. Nat Commun 13 , 2557 (2022). https://doi.org/10.1038/s41467-022-29923-z