


Tarasy rolnicze ( andeny ) były powszechnie budowane i wykorzystywane do celów rolniczych w Andach. Domena publiczna.
Streszczenie
Przez wiele lat rozwój społeczeństw warstwowych wzdłuż środkowego wybrzeża Andów, znanego jako miejsce narodzin cywilizacji andyjskiej, był ściśle powiązany z gospodarką zorientowaną na morze. Hipoteza ta została ostatnio podważona przez rosnące dowody na zarządzanie roślinami i ich uprawę wśród populacji andyjskich na długo przed pojawieniem się złożonych społeczeństw i monumentalnej architektury. Zakres, w jakim gospodarka morska i roślinna były zintegrowane, oraz ich wkład we wczesny osiadły tryb życia, wzrost populacji i stratyfikację wewnątrz społeczności pozostają jednak przedmiotem trwającej i spornej debaty. Korzystając z bayesowskich modeli mieszania ponownie analizujemy wcześniej opublikowane wartości stabilnych izotopów ( kolagen δ 15 N , kolagen δ 13 C , apatyt δ 13 C ) 572 osób z 39 stanowisk archeologicznych w środkowych Andach datowanych na okres od ok. 7000 p.n.e. do 200 n.e., aby zrekonstruować reżimy dietetyczne w kategoriach probabilistycznych. Nasze wyniki wskazują, że ryby, fauna lądowa i rośliny uprawne w zróżnicowany sposób przyczyniały się do diety prehistorycznych populacji andyjskich; w osadach nadmorskich i w dolinach środkowych to uprawa roślin, a nie rybołówstwo, napędzała rozwój najwcześniejszych społeczeństw złożonych w okresie formowania (od 3000 r. p.n.e.). Podobnie, na terenach górskich społeczności, które budowały centra ceremonialne, charakteryzują się gospodarką opartą na roślinach. Nasze odkrycia wskazują również, że kukurydza stała się podstawowym pożywieniem (ponad 25% udziału w diecie) dopiero w nowszych fazach prehistorii andyjskiej, około 500 r. p.n.e.
———————–
W ciągu ostatnich trzech dekad badania archeologiczne zrewolucjonizowały nasze pojmowanie osi czasu i procesów związanych z wczesną udomowieniem roślin wzdłuż środkowego wybrzeża Andów, regionu znanego jako kolebka cywilizacji andyjskiej 1 , 2 , 3 , 4 . Ta transformacja zawdzięcza wiele skrupulatnej analizie mikro- i makroszczątków roślin, które dostarczyły bezcennych spostrzeżeń na temat chronologicznej sekwencji i możliwych dróg, którymi udomowione uprawy zostały rozpowszechnione w całym regionie 5 , 6 , 7 , 8 . Badania wskazują, że w okresie późnego plejstocenu, wczesnego holocenu i środkowego holocenu prehistoryczne populacje w środkowych Andach utrzymywały się dzięki gromadzeniu różnorodnej gamy fauny lądowej i morskiej. Jednocześnie rozpoczęły one zarządzanie i uprawę roślin półudomowionych i udomowionych już w 8350 r. p.n.e. 9 , 10 . Rozwój tych systemów produkcji żywności został uzupełniony rozwojem technologii nawadniających już w 4550 r. p.n.e. 11 oraz udomowieniem wielbłądowatych (między 6500 a 3500 r. p.n.e.) 12 , co doprowadziło do głębokich zmian gospodarczych i społecznych.
Co godne uwagi, kompleksowy przegląd i kalibracja dostępnych datowań radiowęglowych związanych z pozostałościami żywności obejmującymi okres od późnego plejstocenu do okresu Inków ujawniły, że spośród 51 roślin używanych w czasie kontaktu z Europejczykami, co najmniej 40 było uprawianych w epoce przedceramicznej, czyli przed około 1800 r. p.n.e. 12. Ogólnie rzecz biorąc, pojawiające się dowody sugerują, że przerwa czasowa między wczesnym przyjęciem roślin uprawnych a pojawieniem się „pełnoprawnego rolnictwa” (sensu Zeder 13, systemu gospodarczego opartego głównie na produkcji i poleganiu na zwierzętach domowych) mogła nie być tak długa, jak wcześniej zakładano 14 , 15 , co wskazuje, że społeczeństwa przedceramiczne odpowiedzialne za wczesne centra monumentalne prawdopodobnie praktykowały rolnictwo.
Inne linie dowodowe również potwierdzają tę hipotezę. Badania dynamiki demograficznej w Ameryce Południowej przed kontaktem z Ameryką wykazały okres wykładniczego wzrostu populacji, rozpoczynający się około 3000–2000 r. p.n.e., związany ze zmianami środowiskowymi i rozwojem systemów rolniczych. W Andach Środkowych wzrost gęstości osadnictwa sugeruje, że wskaźniki wzrostu populacji były szczególnie wysokie w latach 2000–1 p.n.e. [ 16 ] Jest to zgodne z innymi dowodami empirycznymi na wzrost populacji w okresie formowania (3000–1 p.n.e.), w tym na większe rozmiary i zwartą dystrybucję osad, a także zmiany w architekturze domowej [17 , 18 , 19] .
Jednakże w przypadku wybrzeża peruwiańskiego rola rybołówstwa i rolnictwa w procesach złożoności społecznej, odzwierciedlonej w monumentalnej architekturze i centrach ceremonialnych, hierarchizacji miejsc i stratyfikacji wewnątrzspołecznej, co z kolei może odzwierciedlać istnienie dużej (regionalnej) współpracy społecznej lub kontroli pracy, a być może także istnienie scentralizowanych instytucji 1 , 2 , 15 , 19 , 20, było szeroko dyskutowane 15 , 19 , 20 , 21 , 22 . Przełomowe prace podkreślały, że gospodarka morska była kluczowym czynnikiem rozwoju osiadłego trybu życia, wzrostu populacji i stratyfikacji społecznej w regionie, sugerując, że podziały społeczne powstały wokół wczesnej centralizacji i strategicznej dystrybucji produkcji morskiej, co mogło „wstępnie przystosować” grupy przybrzeżne do późniejszego, coraz bardziej hierarchicznego rozwoju opartego na stosunkowo późnym rolnictwie. W tym scenariuszu rolnictwo wprowadzono długo po początkowym okresie złożoności, około 1800 r. p.n.e., jako sposób na utrzymanie wcześniejszych zinstytucjonalizowanych struktur społecznych 15 .
Późniejsze odkrycia starszych i jeszcze większych stanowisk monumentalnych w środkowych dolinach wzmocniły jednak hipotezę, że cywilizacja powstała w głębi lądu w oparciu o gospodarkę rolną, a nie na wybrzeżu. Co najmniej 35 zaplanowanych stanowisk zintegrowanych z hierarchicznymi systemami wewnątrz dolin na północno-środkowym wybrzeżu Peru, z ponad 200 datowaniami radiowęglowymi między 4100–1800 r. p.n.e., skłoniło archeologów do ponownego przemyślenia początków procesów urbanizacji i ponownego zbadania chronologii Centralnych Andów jako całości1 , 23. Ponadto dowody w postaci pozostałości makro- i mikrobotanicznych pokazują, że gospodarki przybrzeżne były bardziej zróżnicowane i bogatsze w udomowione rośliny, niż wcześniej sądzono24 , 25 , 26 . Nawet w modelach, które minimalizują czynniki materialne jako „wyzwalacze” stratyfikacji społecznej i kładą nacisk na przyczyny behawioralne, takie jak strategie gromadzenia władzy i manipulowanie systemami wierzeń, istnienie systemów utrzymania zdolnych do utrzymania rosnącej populacji w osadach osiadłych odgrywałoby kluczową rolę 27 , 28 .
Zgodnie z ostatnimi odkryciami, od co najmniej okresu formowania początkowego (3000–1 p.n.e.) społeczności dolin basenu Pacyfiku były zorganizowane wokół symbiotycznej gospodarki rolno-rybackiej, z sieciami handlowymi wzdłuż poprzecznych korytarzy litoralnych i śródlądowych, które zwykle nie przekraczały 80–100 km 29 . Społeczeństwa przybrzeżne polegały głównie na rybołówstwie z pewną zależnością od produktów uprawnych, podczas gdy społeczności w środkowych i wyższych sektorach dolin uprawiały półudomowione lub udomowione rośliny uzupełniane produktami morskimi, takimi jak solone na sucho ryby i mięczaki11, 29, 30. Na wyżynach archeologiczne zapisy ośrodków ceremonialnych sugerują, że utrzymanie było w dużej mierze oparte na lądowych produktach żywnościowych31 , 32 , 33. Jednakże względny udział roślin oraz lądowych i morskich źródeł białka w diecie na różnych wysokościach i szerokościach geograficznych nie jest dobrze poznany, a ich znaczenie dla procesu złożoności społecznej pozostaje kontrowersyjne 29 , 30 , 34 , 35.
Ponadto ścieżki intensyfikacji rolniczej niektórych roślin i ich rola w podtrzymywaniu gęstych populacji i struktur społecznych na różnych etapach procesu są nadal przedmiotem dyskusji 26 , 30 , 36. Spośród nich kukurydza ( Zea mays ) jest szczególnie interesująca. Kukurydza rozpoczęła proces udomowienia około 9000 lat temu na nizinach Meksyku 37 i rozprzestrzeniła się do Ameryki Południowej około 6000 r. p.n.e., gdzie ewoluowała w izolacji od dzikiej puli genów teosinte 38 . Dotarła do północnego wybrzeża Peru już w latach 4800–4500 p.n.e. 8 , prawdopodobnie poprzez wielokrotne fale kolonizacji 39 , 40 . Do niedawna Centralne Andy były uważane za „drugie centrum udomowienia kukurydzy” 36 , ale obecnie są uznawane za „centrum udoskonalania”, w którym nastąpiły lokalne adaptacje częściowo udomowionej kukurydzy 38 . Chociaż znaczenie kukurydzy jako podstawowego pożywienia wykazano w późniejszych okresach i powiązano je ze strategiami dominacji państw i imperiów (np. piwo kukurydziane podawane podczas prac społecznych, uczt i ceremonii) 12 , 41 , 42 , jej znaczenie jako siły napędowej najwcześniejszych złożonych społeczeństw pozostaje niejasne30 , 31 , 43 , 44 .
Dostępne dowody wskazują, że jego spożycie mogło być ograniczone w regionie, prawdopodobnie tylko w kontekstach ceremonialnych lub jako uprawa sezonowa, aż do późniejszych czasów 45, 46. Analizy stabilnych izotopów węgla i azotu kolagenu (δ 13 C coll i δ 15 N) i bioapatytu (δ 13 C ap ) z ekshumowanych ludzkich kości i zębów okazały się niezrównaną metodą badania przetrwania w przeszłych populacjach 47. δ 13 C coll jest wskaźnikiem źródeł białka w diecie i szlaków fotosyntezy roślin w sieci pokarmowej (C 3 , C 4 , CAM). δ 13 C ap natomiast odzwierciedla skład izotopowy wszystkich makroskładników (węglowodanów, lipidów, białek) w diecie danej osoby, oferując wgląd w składniki energetyczne 48 . Analiza δ 13 C coll i δ 13 C ap może skutecznie odróżniać diety oparte na C 3 od C 4 (tj. kukurydzy i amarantusa, rośliny wysokogórskiej 12 ) i roślin CAM (kaktusów z wartościami izotopów, które nakładają się na C 4 w przypadku Andów) ze względu na fundamentalne różnice w ich składzie izotopowym 49 , 50 ; jednak nie nadają się one do rozróżniania diet opartych na zasobach morskich od tych opartych na roślinach C 4 ze względu na nakładające się wartości 51 . To ograniczenie jest zwykle rozwiązywane poprzez analizę wartości δ 15 N, ponieważ jest ona obciążona stosunkowo dobrze ugruntowanymi frakcjonowaniami troficznymi (3–6‰), które są większe w systemach wodnych ze względu na dłuższe i bardziej złożone sieci pokarmowe w porównaniu z systemami lądowymi 52 . Wysokie wartości δ 15 N mogą być jednak również związane z suchością 53 i praktykami nawożenia (np. odchodami wielbłądowatych, guanem ptaków morskich) 54 , 55 , a więc dwoma czynnikami, które należy brać pod uwagę podczas analiz izotopowych w Andach Centralnych.
Analizy stabilnych izotopów zostały z powodzeniem zastosowane do rekonstrukcji diety w prehistorycznych populacjach andyjskich, szczególnie w późniejszych okresach, w których zidentyfikowano główne trendy i zmiany dietetyczne17 . Jednakże dowody na wcześniejsze okresy, kiedy powstały pierwsze złożone społeczeństwa, są bardziej ograniczone30 , 34 , 44 , 46. Pomimo doskonałego zachowania morfologicznego szczątków ludzkich i rosnącego zainteresowania związkiem między systemami utrzymania a zmianami społecznymi, wysiłki mające na celu zrozumienie zmian dietetycznych na poziomie jednostki lub populacji zostały utrudnione z powodu braku żywotnego kolagenu z prehistorycznych populacji po pierwszym tysiącleciu n.e.30 , 56 , 57 , 58. Co więcej, do niedawna stosowane metody analityczne pozwalały jedynie na jakościowe podejście do składników diety, które nie pozwalało na ilościowe określenie udziału roślin C3 ( np. bulw, roślin strączkowych) i roślin C4, takich jak kukurydza, ani udziału białek morskich i lądowych w tych wczesnych dietach.
Tutaj, ponownie oceniamy stabilne wartości izotopów węgla i azotu 572 prekolumbijskich osób z Peru. Osoby te reprezentują 61 populacji datowanych na okres od ok. 7000 p.n.e. do 200 n.e., poprzez okres archaiczny (7000–3000 p.n.e.), formatywny (3000–1 p.n.e.) i wczesny okres przejściowy (1–600 p.n.e.), i zostały odzyskane z 39 stanowisk archeologicznych położonych wzdłuż wybrzeża, środkowych dolin i wyżyn trzech odrębnych regionów równoleżnikowych (tj. północy, centrum i południa) Centralnych Andów, charakteryzujących się kontrastującymi warunkami środowiskowymi (Rys. 1 , patrz Informacje uzupełniające 1 ). Uwzględniliśmy również wartości izotopów z 408 okazów fauny i 404 okazów roślinnych rozróżnionych według podregionu, aby scharakteryzować regionalne i lokalne sieci pokarmowe (Informacje uzupełniające 2 ).

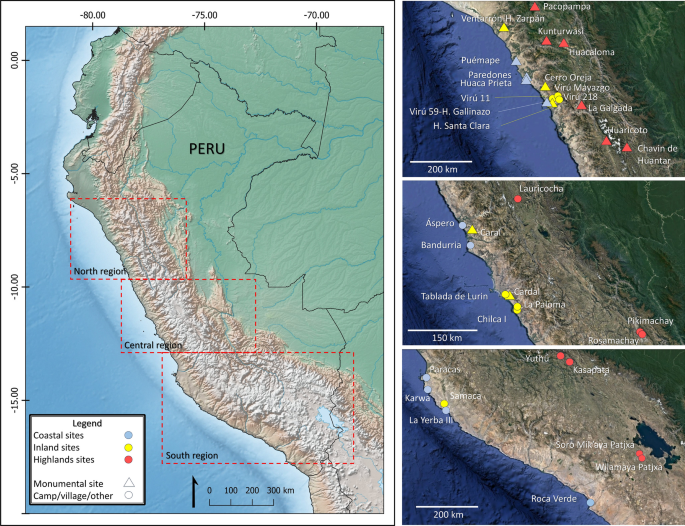
Lokalizacja stanowisk archeologicznych wymienionych w tekście i tabelach. Mapa wygenerowana w programie QGIS w wersji 3.2.8 ( https://www.qgis.org ) z obrazami z Google Earth Pro® (2023 Google LLC. Wszelkie prawa zastrzeżone) oraz rastrową mapą bazową z https://www.naturalearthdata.com/ .
Do oszacowania proporcjonalnego udziału kalorycznego fauny morskiej i lądowej, a także roślin C3 i C4 w diecie osobników z różnych stanowisk i okresów 30, 59, 60, 61 zastosowano podejście bayesowskich modeli mieszania stabilnych izotopów (BSIMM), aby ocenić ich znaczenie gospodarcze dla społeczności andyjskich w okresie archaicznym i formatywnym oraz doprecyzować chronologię głównych zmian w diecie. Spośród 572 osobników, 293 osobniki z pewnymi parametrami zachowania kolagenu62,63 i pełnym zestawem wartości izotopów (tj. δ 13 C coll , δ 15 N i δ 13 C ap ) zostały uwzględnione w rekonstrukcjach BSIMM (patrz Materiały i metody oraz informacje uzupełniające 3 ).
Naszym celem jest ustalenie, czy istotne zmiany społeczne, wzrost demograficzny i przemiany polityczno-ideologiczne, które zazwyczaj definiują periodyzację archeologiczną, są powiązane z intensyfikacją kluczowych upraw, rozumianą jako znaczny wzrost spożycia roślin krajowych. Chociaż naszym głównym celem jest wyjaśnienie roli zasobów morskich i lądowych na wybrzeżu Andów, uwzględniliśmy również diety mieszkańców gór, aby udokumentować różne trajektorie przyjmowania upraw w tym rozległym i heterogenicznym środowisku.
Wyniki
Analiza izotopów stabilnych i rekonstrukcja diety bayesowskiej
W dostępnym zestawie danych wartości δ 13 C coll u osobników ludzkich (n = 317) mieściły się w zakresie od − 23,8‰ do − 9,1‰, wartości δ 15 N (n = 294) od + 2,8‰ do + 24,9‰, a wartości δ 13 C ap (n = 400) od − 15,7‰ do − 2,4‰. Wartości δ 13 C coll i δ 15 N populacji przybrzeżnych mieszczą się w zakresie wartości oczekiwanych dla diet opartych na środowisku morskim, zwłaszcza w okresie archaicznym (7000–3000 p.n.e.), podczas gdy populacje śródlądowe i górskie wykazują wartości zgodne z dietami opartymi na ekosystemach roślinnych C3 ( rys. 2a ). Niektóre populacje wykazują jednak szerokie odchylenia standardowe, co sugeruje dość zróżnicowane diety. Zgodnie z oczekiwaniami wartości δ 15 N u ludzi ulegają znacznemu zmniejszeniu wraz ze wzrostem odległości od morza (n = 292; ρ Spearmana = − 0,693; p < 0,01).

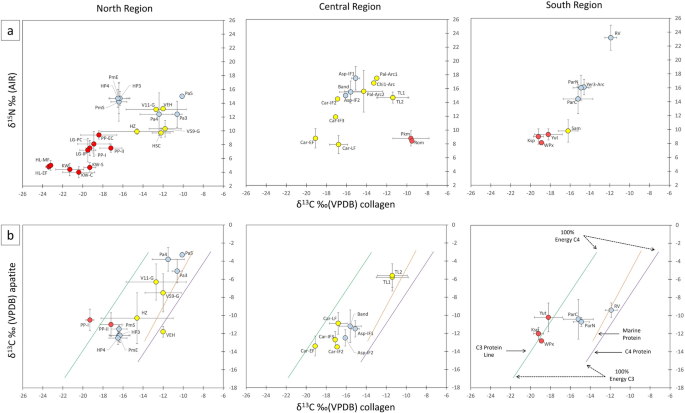
( a ) Średnie wartości δ 13 C coll i δ 15 N z prehistorycznych populacji centralnych Andów według regionu. ( b ) Średnie wartości δ 13 C coll i δ 13 C ap z osobników ludzkich w przestrzeni roboczej Kellnera i Schoeningera 48 , rozróżniane według regionu w celach porównawczych. W tym modelu linie regresji reprezentują źródło białka (C 3 , morskie i C 4 ), a położenie wartości wzdłuż linii wskazuje względny udział energii dostarczanej przez źródło. Czerwone kropki reprezentują populacje górskie. Kropki oznaczają średnie, a krzyżyki odchylenia standardowe (1σ).
Wartości δ 13 C coll i δ 13 C ap (rys. 2 b), które odpowiednio śledzą źródła białka i składników energetycznych diety, pokazują mieszane strategie utrzymania i trzy różne reżimy dietetyczne na przestrzeni czasu: 1) diety populacji początkowej, wczesnoformatywnej i środkowoformatywnej (3000–600 p.n.e.) ze znaczącym udziałem roślin C3 ( tj. roślin strączkowych, bulw i dyniowatych) i bardziej dyskretnym udziałem roślin C4 ( kukurydzy i potencjalnie szarłatu, w zależności od środowiska ekologicznego); 2) diety populacji późnoformatywnej (600–1 p.n.e.) oparte na roślinach C3 ze wzrastającym spożyciem C4 ; i 3) diety oparte na większym spożyciu roślin C4 i mniejszym spożyciu roślin C3 , w większości powiązane z populacjami datowanymi między 200 p.n.e. a pierwszymi wiekami obecnej ery.
Warto zauważyć, że chociaż spożycie roślin C4 znacznie się różniło wśród wczesnych populacji (na przykład wczesna zależność od roślin C4 wydaje się być widoczna na Północnym Wybrzeżu od co najmniej 4500 r. p.n.e. 44 ), ich spożycie znacznie wzrosło w późniejszych populacjach Północnego i Centralnego Wybrzeża, szczególnie w drugiej połowie pierwszego tysiąclecia p.n.e. ( p < 0,05 z Kruskalem–Wallisem-H dla δ 13 C ap ; patrz testy post-hoc w Informacjach uzupełniających 1 : SI1 T4).
Podsumowując, szacunki z BSIMM wskazują, że ogólnie rzecz biorąc, rośliny C3 były głównym źródłem kalorii w diecie dla większości populacji w okresach formowania, zwłaszcza na wyżynach. Zasoby morskie dostarczały najwięcej białka populacjom przybrzeżnym, zmniejszając się w głębi lądu i na wyżynach. Natomiast kalorie z kukurydzy wzrosły z czasem, zwłaszcza wśród populacji przybrzeżnych i śródlądowych z regionów północnych i centralnych w późnym okresie formowania i wczesnym okresie przejściowym (rys. 3 ; patrz również informacje uzupełniające 4 ). Przyznajemy jednak, że jego interpretacja opiera się głównie na pięciu populacjach odstających od normy, a zatem konieczne są dalsze badania w celu potwierdzenia tej hipotezy.

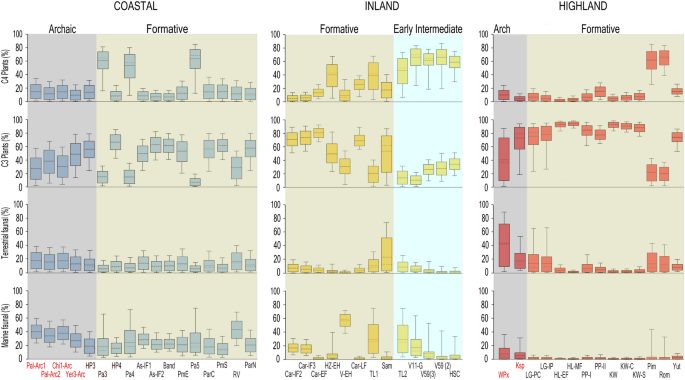
Szacunki kaloryczności diety w odniesieniu do źródeł pożywienia w populacjach andyjskich, pogrupowane według regionów geograficznych i okresów archeologicznych: archaiczny (archaiczny); formacyjny; wczesny pośredni. Pola reprezentują 68% przedział ufności (między 16. a 84. percentylem), a wąsy – 95% przedział ufności (odpowiadający 2,5. a 97,5. percentylowi). Pozioma linia przerywana wskazuje medianę (50. percentyl). Pozioma linia ciągła wskazuje średnią. Identyfikatory stanowisk oznaczone kolorem czarnym odnoszą się do stanowisk lub faz z monumentalną architekturą lub hierarchicznym rozmieszczeniem osad. Identyfikatory oznaczone kolorem czerwonym odnoszą się do stanowisk o nieskomplikowanej strukturze (patrz również SI1 i SI4 F1 i F2).
Dyskusja
Regionalne trendy dietetyczne
W Andach Środkowych rola rolnictwa i rybołówstwa jako czynników napędzających osiadły tryb życia, wzrost populacji i rozwarstwienie społeczne była szeroko dyskutowana, a diety w różnych podregionach wykazują znaczną zmienność w różnych okresach. Populacje zamieszkujące tereny przybrzeżne i górskie wykazują większe uzależnienie od produktów morskich i roślin C3 . Jednak diety mieszkańców terenów przybrzeżnych również wykazują wysokie uzależnienie od roślin C3 . Wśród grup śródlądowych wyraźne nakładanie się wartości izotopowych sugeruje bardziej zróżnicowaną dietę opartą na roślinach.
Na Północnym Wybrzeżu kukurydza była ważnym elementem diety w miejscu Paredones już w latach 4500–4000 p.n.e., podczas gdy w sąsiednim miejscu Huaca Prieta ludzie polegali na żywności morskiej uzupełnianej o produkty lądowe i trochę kukurydzy około 3000–2500 p.n.e. 8 , 10 , 44 , 64. Od wczesnego okresu formacyjnego (1200–1000 p.n.e.) do okresu salinarnego (400–200 p.n.e.) społeczności rozkwitały dzięki połączeniu roślin C3 , białka morskiego i lądowego, z niskim udziałem C4 (<15%) . Badania archeologiczne i analizy koprolitów wskazują na diety złożone z roślin C3 ( tj. bulw, fasoli, orzeszków ziemnych i chleba świętojańskiego), ryb (np. Squatina armata , Mustelus sp., Paralonchurus sp., Cynoscion sp., Sciaena deliciosa ), ssaków morskich ( Otaria sp.) i skorupiaków (np. Mesodesma donacium, Choromytilus chorus, Polinices uber, Thais sp.) oraz pewnej ilości kukurydzy 57 , 65 . Zwiększający się udział kukurydzy, stanowiącej ponad 60% kalorii w diecie, jest wyraźny w stanowiskach śródlądowych (Viru 11, Viru 59) około 200 p.n.e.–200 n.e., w okresie Gallinazo, między późnym okresem formatywnym a wczesnym okresem pośrednim 57 , 66 .
Na północnych wyżynach w ocenianych okresach w dietach dominowały rośliny C3 ( rys. 3 ). Ten scenariusz preferencji C3 ( tj. komosa ryżowa, fasola i różne bulwy) można rozszerzyć regionalnie na inne stanowiska formacyjne z monumentalną architekturą, takie jak Huaricoto (~ 2200–200 p.n.e.), Huacaloma (1500–550 p.n.e.), Pacopampa (900–600 p.n.e.), Chavin (850–450 p.n.e.) i Kuntur Wasi (800–1 p.n.e.). Kukurydza nieznacznie wzrasta tylko na początku obecnej ery31 , 32 , 33 , 46 , 56. Białko lądowe miało potencjalnie większe znaczenie we wczesnych stanowiskach, takich jak La Galgada, prawdopodobnie ze względu na łatwy dostęp jeleniowatych lub wielbłądowatych32 . Później diety były ubogie w białko, pomimo pasterstwa, jak w Pocopampa33 .
Na Wybrzeżu Centralnym populacje archaiczne z La Paloma i Chilca I wykazują największą zależność od fauny morskiej i lądowej spośród wszystkich porównywanych grup34 , 67 (ryc. 3 ) . Chociaż szczątki makro- i mikrobotaniczne (fitolity, skrobia i pyłek) z dolin Fortaleza, Pativilca i Supe datowane na lata 2900–2100 p.n.e. potwierdzają spożycie kukurydzy w okresie formowania początkowego6 , 25 , 26 , analizy izotopowe sugerują , że jej spożycie było ograniczone i sporadyczne, a stanowiska formowania początkowego wykazują zależność od roślin C3 . Na przykład, osoby Caral w okresie formowania początkowego wykazywały udział kalorii w diecie na poziomie 62% roślin C 3 i mniej niż 7% kukurydzy, podczas gdy diety z Áspero i Bandurria30, 68, dwóch wczesnych osad z monumentalną architekturą położonych na linii brzegowej, miały udział 50–60% roślin C 3. Diety roślin C 3 dominowały we wczesnym i środkowym okresie formowania, ale preferencje te zmieniły się w późnym okresie formowania, gdy udział kukurydzy wzrósł o 10% do 25% 30. Na przykład w Tablada de Lurin (200 p.n.e.–100 n.e.), w oparciu o połączenie roślin C 4 i zasobów morskich, kukurydza dostarczała około 40–50% kalorii w diecie 56 , 58.
W regionie południowym, w okresie późnego formowania, osoby ze społeczności Paracas (550–100 p.n.e.), miały dietę składającą się z roślin C3 ( 56–57% kalorii w diecie), głównie bulw (maniok, jíquima, achira , między innymi 69 ), zasobów morskich (19–21% kalorii w diecie) i kukurydzy (11–14% kalorii w diecie). Osoby z cmentarzysk Samaca (100 p.n.e.–100 n.e.) w dolnej dolinie Ica wykazują preferencje dietetyczne wobec roślin C3 ( tj. orzeszków ziemnych, dyni, manioku i pacay ) i niewielkie ilości roślin C4 ( tj. kukurydzy i prawdopodobnie roślin CAM), a także ograniczone pokarmy morskie pomimo stosunkowo niewielkiej odległości od morza 70 . W tym regionie intensyfikacja uprawy kukurydzy wydaje się być opóźniona, pomimo bliskości Ayacucho i Cusco w Południowych Wyżynach, gdzie uprawa kukurydzy była dominująca co najmniej od 800 r. p.n.e. 71 , 72 .
Zebrane dane izotopowe ujawniają trendy dietetyczne w okresie archaicznym (7000–3000 p.n.e.), pokazując, że osoby zamieszkujące tereny przybrzeżne miały zróżnicowaną dietę składającą się głównie z zasobów morskich i roślin C3 ( np. bulw, dyniowatych i roślin strączkowych), obok ssaków lądowych, co jest zgodne ze szczątkami fauny30 , 67. Natomiast osoby zamieszkujące tereny górskie w tym okresie polegały głównie na roślinach C3 i ssakach lądowych. Wkraczając w okres formowania (3000–1 p.n.e.), populacje przybrzeżne, śródlądowe i górskie coraz częściej włączały rośliny C3 do swojej diety, chociaż społeczności przybrzeżne nadal spożywały ryby. Kukurydza odgrywała ograniczoną rolę w okresie formowania, z godnymi uwagi wyjątkami, takimi jak Paredones. Konsumpcja kukurydzy wyraźnie wzrosła w pierwszym tysiącleciu p.n.e., okresie związanym ze wzrostem presji demograficznej16 , 17 , 30 , przyczyniając się do około 25% kalorii w niektórych miejscach śródlądowych do 500 r. p.n.e. Jej intensyfikacja była bardziej wyraźna na północnym wybrzeżu (między 400–100 r. p.n.e.) i wybrzeżu centralnym (~ 200–1 r. p.n.e.), a wolniejsza na południowym wybrzeżu (między 750–450 r. p.n.e.) 73 . Ostatecznie stała się podstawą między 200 r. p.n.e. a 200 r. n.e. na północnym i centralnym wybrzeżu. Ten wzrost produkcji i spożycia kukurydzy w drugiej połowie pierwszego tysiąclecia p.n.e. jest prawdopodobnie związany z produkcją chicha do celów ceremonialnych74 i jest prekursorem ekstensywnej produkcji i wykorzystania chicha w kontekstach wczesnego okresu przejściowego i środkowego horyzontu75 .
Chociaż dowody archeologiczne i botaniczne zdecydowanie potwierdzają, że kukurydza jest główną rośliną C4 w Andach, należy zauważyć, że nie była ona wyłącznym źródłem wartości wzbogaconych w 13C w diecie mieszkańców Andów. Nakładające się wartości δ13C w kukurydzy, szarłacie i roślinach CAM (tj. kaktusach) stanowią czynnik mylący, który należy uwzględnić w interpretacji adopcji kukurydzy w środowiskach suchych i wysokogórskich. Dostępne dowody sugerują jednak, że kaktusy były ważniejsze dla populacji archaicznych, a szarłat, jako podstawowy składnik pożywienia, prawdopodobnie występował wyłącznie na wyżynach lub na niższych szerokościach geograficznych [12 , 70 , 76] .
Ten rozszerzony proces intensyfikacji rolnictwa wśród społeczności andyjskich obejmował wdrożenie produktywnych strategii, które były dobrze dostosowane do konkretnych upraw, gleb i ograniczonych zasobów wodnych w tych suchych i wymagających krajobrazach (tj. zarządzanie wodą podziemną, rosnąca technologia nawadniania, zapadnięte pola, rolnictwo zalewowe, uprawa konsorcyjna itp. 77 ), modulowane przez zmieniające się warunki środowiskowe 16 , 78 , 79 , 80 . Te ulepszenia technologiczne prawdopodobnie nie nastąpiły w tym samym czasie i niekoniecznie były związane z tymi samymi czynnikami w każdym podregionie. Struktury społeczne i polityczne mogły w różny sposób wpływać na sposób, w jaki uprawy były przyjmowane i wykorzystywane przez populacje z różnych wysokości, charakteryzując określone procesy dla każdego regionu i miejsca.
Czynniki, które warunkowały intensyfikację uprawy kukurydzy przez kilka populacji „zależnych od kukurydzy” i uniemożliwiały jej przyjęcie przez większość społeczności formacyjnych, są prawdopodobnie związane z uwarunkowaniami ekologicznymi (np. dostępnością wody, ukształtowaniem terenu, klimatem i ograniczeniami wysokościowymi) oraz kulturowymi (np. tożsamościowymi). Na przykład szersze doliny północnego wybrzeża, zwłaszcza te ze stałym zaopatrzeniem w wodę, są bardziej odpowiednie do uprawy kukurydzy przy użyciu technologii intencjonalnej20 .
Podsumowując, niniejsza analiza pomaga odpowiedzieć na trzy fundamentalne pytania dotyczące diety w okresie archaicznym i formacyjnym środkowych Andów: 1) dieta, która doprowadziła do rozwoju społeczeństw związanych z monumentalną architekturą, prawdopodobnie społeczeństw złożonych, nie była w przeważającej mierze morska pod względem kaloryczności; 2) rozwój ten opierał się głównie na roślinach C3 na każdej wysokości, w tym na obszarach przybrzeżnych; oraz 3) większość diet nie opierała się na kukurydzy, która z kolei była uprawą o marginalnym znaczeniu, przynajmniej do pierwszego tysiąclecia p.n.e. Zatem, zgodnie z wcześniejszymi argumentami, w przypadku obszarów przybrzeżnych, nasze wyniki podważają dominującą teorię, że zasoby morskie były głównym motorem ekonomicznym złożoności społecznej. Nasze wyniki natomiast dobrze wpisują się w wyłaniający się pogląd, że rozwój polityczny w większości obszarów śródlądowych i górskich opierał się na systemach produkcyjnych opartych na uprawach C3 . Podkreśla to znaczenie uprawy roślin w napędzaniu wzrostu populacji, stratyfikacji społecznej i rozwoju instytucji politycznych w jednej z kolebek cywilizacji.
Metody
Stanowiska archeologiczne i osoby
To badanie in silico opracowane z wykorzystaniem wcześniej opublikowanych danych dotyczących stabilnych izotopów, przechowywanych w bazie danych South American Archeological Isotopic Database (SAAID_V.2.0_2023, hostowanej na platformie CORA — https://doi.org/10.34810/data602 ). W naszej analizie uwzględniono łącznie 572 osoby z 39 stanowisk archeologicznych w centralnych Andach, które reprezentują całość dostępnych danych sprzed obecnej ery.
Baza danych z surowymi wartościami izotopów osobników (wartości konsumentów) jest podana w Informacjach uzupełniających 1. Szacunki płci i wieku są podane jak w oryginalnych odniesieniach. Większość stanowisk znajduje się w peruwiańskich dolinach, które poprzecznie przecinają Pustynię Nadbrzeżną Pacyfiku, aby odprowadzić wodę do Oceanu Spokojnego, w lokalizacjach przybrzeżnych (tj. mniej niż 3 km od morza; n = 10) i śródlądowych (tj. dolne i środkowe doliny < 600 m n.p.m., położone w dowolnej odległości od wybrzeża; n = 16). Inne stanowiska (n = 13) znajdują się na wyżynach andyjskich (tj. górne doliny basenu Pacyfiku o wysokości > 600 m n.p.m. oraz doliny międzyandejskie na dowolnej wysokości). Ponieważ niektóre stanowiska mają długie sekwencje zawodowe, przeanalizowano łącznie 61 odrębnych populacji (patrz Informacje uzupełniające 1 ). Stanowiska i fazy sklasyfikowano według złożoności społecznej w dwóch kategoriach: 1) stanowiska z monumentalną architekturą lub centrami ceremonialnymi, które odpowiadają mniej lub bardziej złożonym społeczeństwom; oraz 2) inne typy stanowisk (obozy, miasta, stanowiska otwarte, schroniska skalne), które prawdopodobnie odpowiadają społeczeństwom nieskomplikowanym lub egalitarnym. Klasyfikację tę zestawiono z interpretacjami archeologicznymi dla każdego stanowiska, opartymi na kilku markerach (tj. cechach architektonicznych, skali zabudowy, wzorcach osadnictwa wewnątrz doliny, dowodach grobowych i inwentarzach archeologicznych), a w razie potrzeby skorygowano (np. stanowiska na Półwyspie Paracas, które są grobowcami powiązanymi ze złożoną siecią osadniczą i bogatymi darami grobowymi).
Ramy chronologiczne stanowisk i faz kulturowych oparto na ponownie skalibrowanych datach radiowęglowych podanych w literaturze (szczegóły w Informacjach uzupełniających 3 ). Okres badanych osobników obejmuje okres archaiczny (7000–3000 p.n.e.), formacyjny (3000–1 p.n.e.) i wczesny przejściowy (1–600 p.n.e.).
Nasza analiza objęła również wartości izotopów z 812 okazów fauny (n = 408) i roślin (n = 404) do rekonstrukcji sieci pokarmowej (patrz Informacje Dodatkowe 2 ). Wartości te zostały rozróżnione według regionu szerokości geograficznej, aby uzyskać bardziej wiarygodne dane wejściowe dla modeli bayesowskich i interpretacji wizualnych. Izotopowa linia bazowa potencjalnych źródeł pokarmu dla każdego podregionu została wygenerowana za pomocą rKin przy użyciu Standard Ellipse Areas81 (patrz Informacje Dodatkowe 3 ). Wszystkie wartości izotopów zostały wyodrębnione z SAAID.
Aby zapobiec potencjalnie mylącemu efektowi problemów z tafonomią w naszej analizie, uwzględniono tylko osoby z wiarygodnymi wartościami δ 13 C coll i δ 15 N z kości i kolagenu zębiny, zgodnie z przyjętymi kryteriami konserwacji62, 63. W przypadku wartości δ 13 C ap , chociaż objętość opublikowanych danych jest większa niż dla kolagenu, kryteria konserwacji nie były szeroko stosowane, szczególnie w starszych badaniach, a założenie wiarygodności opierało się w dużej mierze na protokołach ekstrakcji82. W tym badaniu użyliśmy wartości δ 13 C ap kości tylko wtedy, gdy kolagen w próbce był dobrze zachowany. Założyliśmy, że jeśli kolagen, organiczna biocząsteczka, która tworzy strukturę biologiczną mieszczącą kryształy nieorganiczne, jest dobrze zachowany, to bioapatyt również jest dobrze zachowany. Jednak poza spójnością danych (wartości w apatycie są zazwyczaj wzbogacone w 13 C w porównaniu z kolagenem) nie ma innych obiektywnych markerów do oceny zachowania apatytu83. Ponieważ szkliwo zębów zachowuje się lepiej niż kość, wykorzystano wszystkie dostępne wartości δ 13 C ap szkliwa pochodzące od wybranych osób.
Aby uniknąć czynników zakłócających związanych z karmieniem piersią i odstawianiem od piersi, w naszych analizach uwzględniono wyłącznie osoby dorosłe o akceptowalnym składzie atomowym kolagenu. Niemowlęta i małe dzieci (młode dorośli <4 lat) nie zostały uwzględnione84 . Zgodnie z tymi kryteriami, do analiz BSIMM wybrano 293 osoby z pewnymi parametrami zachowania kolagenu i pełnym zestawem wartości izotopowych (tj. δ 13 C coll , δ 15 N i δ 13 C ap ).
Analiza izotopowa i BSIMM
Najpierw użyliśmy statystyki opisowej i konwencjonalnych wykresów rozrzutu izotopów jako narzędzi diagnostycznych do identyfikacji głównych trendów i zmian dietetycznych dla każdej populacji i regionu. Następnie dla każdej grupy przeprowadzono BSIMM. Wykresy rozrzutu δ 15 N vs. δ 13 C coll dostarczają informacji o rodzaju spożywanego białka, lądowym (azot poniżej +11 ‰ i węgiel poniżej -12,5 ‰) lub morskim, jeśli obie wartości są bardzo wysokie 52 . Wykresy rozrzutu δ 13 C coll vs. δ 13 C ap w obszarze roboczym trzech linii regresji uzyskanych z eksperymentalnych badań żywieniowych umożliwiają rozróżnienie źródeł białka i węglowodanów (C 3 lub C 4 ). Rozkład wartości dla danej osoby na osi apatytu wskazuje, czy energia w diecie to C3 czy C4 , a na osi kolagenu, czy źródłem białka jest C3 czy C4 / pochodzenia morskiego. Populacje skoncentrowane między tymi dwiema liniami miałyby złożoną dietę, łączącą białka C3 i C4 . Odległość do linii końcowych wskazuje przybliżony stosunek spożywanej energii z C3 lub C4 48 .
Do wykrycia różnic diagnostycznych w czasie i przestrzeni wykorzystano statystyki opisowe (średnia, zakres, odchylenie standardowe, błąd standardowy średniej) oraz porównania wartości izotopów między grupami. Analizy te przeprowadzono z użyciem testów Kruskala-Wallisa (α = 0,05), po uprzednim sprawdzeniu rozkładu normalnego za pomocą testu Kołmogorowa-Smirnowa (α = 0,05). Jako test post hoc zastosowano test U Manna-Whitneya (α = 0,05). Do sprawdzenia zależności między wartościami izotopów a odległością od morza wykorzystano korelację Spearmana. Wszystkie testy przeprowadzono w oprogramowaniu SPSS v.21 (Microsoft®).
Szeroki zakres zmienności dietetycznych w zakresie składu makroskładników, wartości izotopowych spożywanych produktów, frakcjonowania (równoważenia diety dla konsumenta) oraz czynników wpływających na dystrybucję składników odżywczych, a także inne źródła niepewności, często utrudniają dokładne rekonstrukcje diety wyłącznie za pomocą metod graficznych lub liniowych. Z kolei modele BSIMM pozwalają na uwzględnienie wielu wskaźników i źródeł zmienności, umożliwiając bardziej wiarygodne oszacowania składu diety w postaci rozkładów prawdopodobieństwa59 , 60 , 61 .
Do oszacowania proporcjonalnego udziału kalorii różnych źródeł żywności w diecie człowieka, na podstawie wartości stabilnych izotopów jako wskaźników żywieniowych, wykorzystano oprogramowanie Food Reconstruction Using Isotopic Transferred Signals (program beta FRUITS w wersji 2.1.1) 59 . Zastosowano ważony model z czynnikami frakcjonowania (przesunięciami) opartymi na stężeniach składników odżywczych w poszczególnych grupach żywności60 , 61 .
Dane konsumenckie dla poszczególnych osób składają się z ich wartości izotopowych z niepewnością ± 0,5 60 . W przypadku grup zastosowaliśmy średnie wartości izotopów dla każdego okresu/miejsca i błąd standardowy średniej . Po dokładnym przesianiu kryteriów konserwacji kolagenu w bazie danych SAAID, do analizy włączono tylko osoby z ważnymi wartościami izotopów kolagenu z każdego okresu/miejsca. Jeśli było to możliwe, kompletny zestaw wartości δ 13 C coll , δ 15 N coll i δ 13 C ap uwzględniono w modelach trzech proxy. Ponieważ jednak niektóre osoby/populacje miały tylko wartości δ 13 C coll , δ 15 N coll , musieliśmy wdrożyć alternatywne modele z dwoma proxy w celach porównawczych. Modele z dwoma proxy mogą odzwierciedlać główne trendy w składach diet, ale mogą być mniej pewne co do dokładnych proporcji analizowanych źródeł żywności w porównaniu z modelami z trzema proxy 30 , 59 , 60 .
Potencjalne przyswojone źródła pożywienia ( grupy pożywienia ) wraz z ich składem makroskładników ( frakcje pożywienia: masa, białko i energia) zostały pogrupowane w cztery kategorie: fauna lądowa (dostarczająca białka i lipidy), zwana w modelu „TF”; fauna morska (dostarczająca białka i lipidy), zwana „MF”; oraz rośliny C3 i C4 ( dostarczające węglowodany i białka), zwane odpowiednio C3 i C4 30 , 59 , 60 , 61 .
Wartości izotopów z każdej grupy żywności pochodzą z izotopowych badań paleodietetycznych skupionych na dolinach Andów Centralnych sklasyfikowanych w trzech regionach równoleżnikowych: Wybrzeże Centralne i Wyżyny Centralne, Wybrzeże Północne i Wyżyny Północne oraz Wybrzeże Południowe i Wyżyny Południowe. Dla zachowania spójności, każde stanowisko zostało przeanalizowane zgodnie z danymi izotopowymi z gatunków regionalnych lub lokalnych, zwracając szczególną uwagę na inwentaryzacje archeologiczne stanowiska lub wcześniejsze wartości izotopów z lokalnych współczesnych lub archeologicznych próbek. Chociaż implikuje to pominięcie niektórych cennych odniesień, każdy region został scharakteryzowany za pomocą własnych wartości. Jeśli wartości lokalne lub regionalne nie były dostępne, wykorzystano wartości z regionów sąsiednich. Dla porównań z wartościami archeologicznymi, wartości δ 13 C z okazów współczesnych zostały skorygowane o efekt paliw kopalnych, dostosowując wartości o + 1,5 ‰ 85 .
W przypadku fauny morskiej wykorzystano syntezę kilku źródeł, opartą głównie na współczesnych danych z Wybrzeża Centralnego30 . W przypadku roślin wykorzystano wyłącznie wartości izotopów z okazów współczesnych, aby uniknąć problemów związanych z konserwacją86 , 87. Średnie wartości δ13C i δ15N grup pokarmowych, liczbę okazów i gatunków uwzględnionych w modelach, a także oryginalne odniesienie do wartości dla regionów centralnego, północnego i południowego, można znaleźć w Informacjach uzupełniających2.
Współczynniki frakcjonowania izotopów to wartości konsensualne pochodzące z badań eksperymentalnych. Dla δ 13 C współczynnik frakcjonowania ustalono na + 4,8 ± 0,5 ‰ między dietą a kolagenem oraz + 10,1 ± 0,5 ‰ między dietą a apatytem. Współczynnik frakcjonowania diety i kolagenu dla δ 15 N ustalono na + 5,5 ± 0,5 ‰ 59 , 60 , 61 , 88 .
Ważone wartości każdej frakcji makroskładników (lipidów, węglowodanów i białek) ustalono na podstawie wcześniej opublikowanych parametrów 30 , 59 , 60 , 61. Wartości δ 13 C ap reprezentują całkowitą mieszankę węgla w diecie. Dlatego stosujemy tę samą wartość masową dla każdej grupy żywności 56 , 57 , 58. Wartości masowe δ 13 C dla fauny lądowej i morskiej oszacowano jako średnią ważoną wartości lipidów i białek δ 13 C 59 , 60 W przypadku kolagenu węgiel białka i energia kierowana do całkowitego kolagenu ustalono na odpowiednio 74 ± 4% i 26% 89 , chociaż węgiel może być kierowany z węglowodanów i lipidów poddanych recyklingowi podczas syntezy nieistotnych aminokwasów 90. Założyliśmy, że azot pochodził wyłącznie z białek (100%). Lipidy i węglowodany dodano do modelu jako „energię”. Aby zintegrować ten efekt kombinatoryczny, zastosowaliśmy model „zależny od stężenia” 59 , 60 , 61 .
Zgodnie z poprzednimi rekonstrukcjami przy użyciu FRUITS 30 , 59 , 60 , 61 , skład izotopowy każdej frakcji odżywczej (białka, węglowodanów i lipidów) uzyskano ze średnich wartości δ 13 C coll i δ 15 N coll przy użyciu następujących współczynników frakcjonowania: − 2‰ ( ∆ 13 C białko-kolagen ), − 8‰ ( ∆ 13 C lipidy-kolagen ) i + 2‰ (∆ 15 N białko-kolagen ) dla ssaków lądowych i − 1‰ ( ∆ 13 C białko-kolagen ), − 7‰ (∆ 13 C lipidy-kolagen ) i + 2‰ (∆ 15 N białko-kolagen ) dla ssaków morskich zwierzęta. W przypadku roślin przesunięcia wynosiły − 2‰ (∆ 13 C masa-białko ) i + 0,5‰ (∆ 13 C masa-lipidy ), natomiast dla wartości δ 15 N białka roślinnego przyjęto znaną wartość δ 15 N zarejestrowaną dla rośliny. Masę węgla ( stężenia ) każdej frakcji pokarmu (białko i energia) z każdej grupy pokarmów obliczono na podstawie jej składu makroskładników 30 , 59 , 60 , 61 (szczegóły w Informacjach uzupełniających 3 ).
Na koniec szacunki dietetyczne sparametryzowano za pomocą fizjologicznego, konserwatywnego i akceptowalnego zakresu spożycia białka określonego między 5 a 45% całkowitej liczby kalorii59 . Te dane wejściowe zostały obciążone jako „wcześniejsze”. Kierując się naszymi obserwacjami wykresów dwuwymiarowych, przetestowano alternatywne modele dla każdego miejsca i populacji. Szacunki wygenerowane dla modeli FRUITS odzwierciedlają zawartość węgla lub równoważne wkłady kaloryczne i są wyrażone jako wkłady względne (sumujące się do 1 lub 100%) z powiązaną niepewnością 1σ60 . Szacunki wkładów grup żywności przedstawiono w Informacjach uzupełniających 4 (patrz również Informacje uzupełniające 5 ).
Ograniczenia badania
Należy obiektywnie ocenić ograniczenia niniejszego badania i uznać inherentną złożoność procesów gospodarczych i społeczno-kulturowych w regionie andyjskim. Procesy te wykraczają poza to, co można uchwycić wyłącznie za pomocą rekonstrukcji diety. Wraz z pojawianiem się nowych badań, obecne rozumienie tematu może ewoluować, a my musimy pozostać otwarci na rewizję naszych interpretacji w oparciu o dodatkowe ustalenia.
Ponadto badanie to ujawniło pewne czynniki komplikujące, które mogły mieć wpływ na ogólną interpretację danych: 1) informacje pozwalające ocenić zachowanie kolagenu kostnego zgodnie z ustalonymi kryteriami jakości kolagenu 62 , 63 nie zawsze były dostępne w badanej literaturze; 2) kilka starożytnych populacji andyjskich charakteryzowało się słabym zachowaniem kolagenu kostnego, co nie zapewniało wystarczającej ilości kolagenu kostnego do datowania lub analiz izotopowych, a w danych izotopowych z próbek zaobserwowano znaczne luki czasowe i regionalne; 3) miejsca te zawierały różną liczbę osób, co może mieć wpływ na obserwowaną zmienność; 4) w niektórych miejscach brakowało informacji na temat szczątków fauny i botanicznych, a dane uniemożliwiały ustalenie lokalnych baz izotopowych, co ograniczyło możliwość uzyskania dokładniejszych szacunków dietetycznych; 5) większość badań z ostatnich trzech dekad uwzględnionych w tym badaniu nie miała dostępu do metod obecnie stosowanych do analizy integralności bioapatytu (tj. spektroskopii Ramana i FTIR) 83 , dlatego nie można oszacować diagenezy szkliwa i bioapatytu; ponadto istnieje potencjalny problem ze zmiennością międzylaboratoryjną analizy δ13Cap91 ; 7 ) pomimo dokładności zastosowanych metod, powinniśmy wziąć pod uwagę ekwifinalność izotopową, czyli różne kombinacje wkładów żywnościowych , które mogą skutkować takimi samymi wartościami izotopowymi dla konsumenta; wreszcie, 8) markery izotopowe nie są w stanie odróżnić roślin uprawnych od nieuprawianych, co jest kluczowe w tej dyskusji; jednak na podstawie dostępnych odniesień2 , 4 , 7 , 10 , 12 , 26 możemy założyć, że większość roślin, których wartości izotopowe zaobserwowano w tym badaniu, to uprawy udomowione.
Dostępność danych
Wszystkie dane potwierdzające wnioski z artykułu znajdują się w tekście głównym oraz w plikach z informacjami uzupełniającymi. Dane dotyczące izotopów stabilnych wykorzystane w niniejszym badaniu zostały wcześniej opublikowane i uzyskane zgodnie z przepisami rządu peruwiańskiego (tj. Ministerio de Cultura od 2010 roku, a wcześniej Instituto Nacional de Cultura ). Badania te nie obejmowały bezpośredniego przetwarzania tkanek ludzkich i nie wiążą się z żadnymi problemami etycznymi związanymi z przetwarzaniem opublikowanych danych.
Odniesienia
-
Shady Solís, R., Haas, J. i Creamer, W. Dating Caral, preceramiczny ośrodek miejski w dolinie Supe na środkowym wybrzeżu Peru. Science 292 , 723–726 (2001).
-
Haas, J. i Creamer, W. Tygiel cywilizacji andyjskiej: wybrzeże peruwiańskie od 3000 do 1800 r. p.n.e. Curr. Anthropol. 47 , 745–756 (2006).
-
Dillehay, TD Gdzie ląd spotyka się z morzem: czternaście tysiącleci historii ludzkości w Huaca Prieta, Peru (University of Texas Press, 2017).
-
Dillehay, TD Od zbieractwa do rolnictwa w Andach: nowe perspektywy produkcji żywności i organizacji społecznej (Cambridge University Press, 2011).
-
Grobman, A. i in. Kukurydza preceramiczna firm Paredones i Huaca Prieta. Peru. Proc. Natl. Acad. Nauka. USA 109 , 1755–1759 (2012).
-
Haas, J. i in. Dowody na obecność kukurydzy ( Zea mays ) w późnym archaiku (3000–1800 p.n.e.) w regionie Norte Chico w Peru. Proc. Natl. Acad. Sci. USA 110 , 4945–4949 (2013).
-
Bonavia, D. i in. Szczątki roślin w książce Where the Land Meets the Sea: Fourteen Millennia of Human History at Huaca Prieta, Peru (red. Dillehay, TD) 367–433 (University of Texas Press, Austin, TX, 2017).
-
Prieto, G. Ogród rybaka: praktyki ogrodnicze w morskiej społeczności drugiego tysiąclecia na północnym wybrzeżu Peru w: Maritime Communities of the Ancient Andes (red. Prieto, G. i Sandweiss, DH) 218–246 (University Press of Florida, 2020).
-
Dillehay, TD, Rossen, J., Andres, TC i Williams, DE Wstępne zastosowanie orzeszków ziemnych, dyni i bawełny w północnym Peru. Science 316 , 1890–1893 (2007).
-
Piperno, DR i Dillehay, TD Ziarna skrobi na zębach ludzkich ujawniają wczesną dietę szeroko rozpowszechnioną w północnym Peru. Proc. Natl. Acad. Sci. USA 105 , 19622–19627 (2008).
-
Dillehay, TD, Elling, H. i Rossen, J. Preceramiczne kanały irygacyjne w peruwiańskich Andach. Proc. Natl. Acad. Sci. USA 102 , 17241–17244 (2005).
-
León, E. 14,000 años de alimentación en el Perú (Universidad de San Martin de Porres, 2013).
-
Zeder, MA Podstawowe pytania w badaniach nad udomowieniem. Proc. Natl. Acad. Sci. USA 112 , 3191–3198 (2015).
-
Dillehay, TD i in. Wyłaniająca się konsiliencja wśród współczesnych społeczności rybackich i rolniczych środkowego holocenu na północnym wybrzeżu Peru. Front. Earth Sci. 10 , 939214 (2022).
-
Moseley, ME Morskie podstawy cywilizacji andyjskiej (Cummings, Menlo Park, CA, 1975).
-
Goldberg, A., Mychajliw, AM i Hadly, EA. Demografia ludzi prehistorycznych w Ameryce Południowej po inwazji. Nature 532 , 232–235 (2016).
-
Wilson, KM i in. Klimat i demografia napędzają 7000 lat zmian w diecie mieszkańców Andów Centralnych. Sci. Rep. 12 , 2026 (2022).
-
Willey, G. Prehistoryczne wzorce osadnictwa w dolinie Virú w Peru (Bureau of American Ethnology Bulletin 155, Smithsonian Institution, Waszyngton DC, 1953).
-
Sandweiss, DH Wczesne rybołówstwo i zabytki śródlądowe. W cywilizacji andyjskiej. Hołd dla Michaela Moseleya (red. Marcus, J. i Williams, PR) 39–54 (Cotsen Institute of Archaeology, Uniwersytet Kalifornijski, Los Angeles, Kalifornia, 2009).
-
Engel, FA De las Begonias al Maíz. Vida y Producción en el Perú Antiguo (Centro de Investigaciones de Zonas Áridas CIZA, Universidad Nacional Agraria La Molina, Lima, 1987).
-
Raymond, JS Morskie podstawy cywilizacji andyjskiej: rekonstrukcja dowodów. Am. Antiq. 46 , 806–821 (1981).
-
Makowski, K. Prehiszpański andyjski urbanizm i jego „antymiejskie” osobliwości. J. Urban Archaeol. 8 , 165–196 (2023).
-
Creamer, W., Ruiz, A., Perales, M. i Haas, J. Dolina Fortalezy, Peru: badania archeologiczne stanowisk późnoarchaicznych (3000–1800 p.n.e.). Fieldiana Anthropol. 44 , 1–108 (2013).
-
Vega-Centeno, R. Rytuał i architektura w kontekście wyłaniającej się złożoności: perspektywa z Cerro Lampay, późnoarchaicznego stanowiska w środkowych Andach (rozprawa doktorska, Uniwersytet Arizony, Tucson, AZ, 2005).
-
Shady, R. Los orígenes de la Civilización y la formación del Estado en el Perú: Las evidencias arqueológicas de Caral-Supe. W La Ciudad Sagrada de Caral-Supe: Los orígenes de la Civilización andina y la formación del Estado Prístino en el antiguo Perú (red. Shady, R. i Leyva, L.) 93–105 (Instituto Nacional de Cultura, Lima, 2003).
-
Yseki, M., Pezo-Lanfranco, L., Machacuay, M., Novoa, P. i Shady, R. Spożywanie roślin w Áspero w Peru w początkowym okresie formowania (3000–1800 p.n.e.): nowe dowody z ziarna skrobi uwięzionego w ludzkim kamieniu nazębnym. Sci. Rep. 13 , 14143 (2023).
-
Rick, JW, Ewolucja autorytetu i władzy w Chavín de Huántar, Peru. W: Foundations of Power in the Prehispanic Andes (red. Vaughn, KJ, Ogburn, D. i Conlee, Kalifornia), s. 13–36 (Archeological Papers of the American Anthropological Association, tom 14, Arlington, Wirginia, 2005).
-
Moore, J. Władza i praktyka w Andach Prehispanic: Uwagi końcowe. W: „ Fundamenty władzy w Andach Prehispanic” (red. Vaughn, KJ, Ogburn, D. i Conlee, Kalifornia) 261–274 (Archeological Papers of the American Anthropological Association, tom 14, Arlington, Wirginia, 2005).
-
Shady, R. El sistema social de Caral y su trascendencia: El manejo transversal del territorio; la complementaredad social i politica; y la interacción intercultural, Nayra Kunan Pacha, 19–90 (2018).
-
Pezo-Lanfranco, L. i in. Dieta na początku cywilizacji andyjskiej: nowe dane dotyczące stabilnych izotopów z Caral i Áspero na północno-środkowym wybrzeżu Peru. Jestem. J. Biol. Antropol. 177 , 402–424 (2022).
-
Seki, Y. i Yoneda, M. Cambios de manejo del poder en el Formativo: desde el análisis de la dieta alimenticia. Perspectivas Latinoamericanas 2 , 110–131 (2005).
-
Washburn, E. i in. Kukurydza i zmiany w diecie we wczesnej cywilizacji peruwiańskiej: dowody izotopowe ze stanowiska La Galgada w Peru z późnego okresu preceramicznego/okresu inicjalnego. J. Arch. Sci. Rep. 31 , 102309 (2020).
-
Takigami, M. i in. Badanie izotopowe eksploatacji kukurydzy w okresie formowania w Pacopampie. Peru. Anthropol. Sci. 129 (2), 121–132 (2021).
-
Beresford-Jones, DG i in. Dieta i styl życia w pierwszych wioskach środkowego preceramiku: wnioski z analiz stabilnych izotopów i osteologicznych szczątków ludzkich z Palomy, Chilca I, La Yerba III i Morro I. Lat. Am. Antiq. 32 , 741–759 (2021).
-
Beresford-Jones, DG i in. Doskonalenie morskich fundamentów cywilizacji andyjskiej: Jak technologia włókien roślinnych napędzała złożoność społeczną w okresie przedceramicznym. J. Arch. Method Theory 25 , 393–425 (2018).
-
Bonavia, DE maíz, su origen, su domación y el rol que ha cumplido en el desarrollo de la Culture (Universidad de San Martín de Porras, 2008).
-
Piperno, DR, Ranere, AJ, Holst, I., Iriarte, J. i Dickau. R. Proc. Natl. Acad. Nauka. USA 106 , 5019–5024 (2009).
-
Kistler, L. i in. Dowody wieloprocesorowe wskazują na złożoną historię ewolucyjną kukurydzy w Ameryce Południowej. Science 362 , 1309–1313 (2018).
-
Moreno-Mayar, JV i in., Wczesne rozprzestrzenianie się ludzi w Ameryce. Science 362 , eaav2621 (2018).
-
Nakatsuka, N. i in. Rekonstrukcja paleogenomiczna głębokiej historii populacji Andów. Cell 181 , 1–15 (2020).
-
Rostworowski, M. Pescadores, artesanos y mercaderes costeños en el Perú prehispánico w Obras Completas III (Instituto de Estudios Peruanos IEP, Lima, 2004).
-
Tung, TA i Knudson, KJ Analiza stabilnych izotopów przedhiszpańskiej społeczności andyjskiej: rekonstrukcja diety przedwojennej i ery Wari w głębi imperium Wari w Peru. Jestem. J.Fiz. Antropol. 165 , 149–172 (2018).
-
Perry, L. i in. Wczesna uprawa kukurydzy i interakcje międzystrefowe w południowym Peru. Nature 440 , 776–779 (2006).
-
Tung, TA, Dillehay, TD, Feranec, RS i DeSantis, LRG Wczesne wyspecjalizowane gospodarki morskie i kukurydziane na północnym wybrzeżu Peru. Proc. Natl. Acad. Sci. USA 117 , 32308–32319 (2020).
-
Shady, R. Caral-Supe i północno-centralny obszar Peru: Historia kukurydzy na ziemi, na której narodziła się cywilizacja, w: Historie kukurydzy: Interdyscyplinarne podejścia do prehistorii, językoznawstwa, biogeografii, udomowienia i ewolucji kukurydzy (red. Staller, JE, Tykot, RH i Benz, BF) 381–402 (Elsevier, Nowy Jork, 2006).
-
Burger, R. & van der Merwe, N. Kukurydza i pochodzenie cywilizacji górskiej Chavín: perspektywa izotopowa. Am. Anthropol. 92 , 85–95 (1990).
-
Richards, M. Analiza izotopów w badaniach nad dietą. W: Archaeological Science: An Introduction (red. Richards, M. i Britton, K.) 125–144 (Cambridge University Press, Cambridge, 2020).
-
Kellner, CM i Schoeninger, MJ Prosty model izotopu węgla do rekonstrukcji diety człowieka prehistorycznego. Am. J. Phys. Anthropol. 133 , 1112–1127 (2007).
-
Ambrose, SH i Norr, L. Dowody eksperymentalne na związek między stosunkami izotopów węgla w diecie pełnoporcjowej i białku dietetycznym a stosunkami kolagenu i węglanu kostnego w prehistorycznych kościach człowieka: archeologia na poziomie molekularnym (red. Lambert, J. i Grupe, G.) 1–37 (Springer, Berlin, 1993).
-
Lee-Thorp, JA, Sealy, JC i van der Merwe, NJ Różnice w stosunkach stabilnych izotopów węgla pomiędzy kolagenem kostnym a apatytem kostnym oraz ich związek z dietą. J. Archaeol. Sci. 16 , 585–599 (1989).
-
Chisholm, BS Zmienność rekonstrukcji diety na podstawie dowodów w postaci stabilnych izotopów węgla w The Chemistry of Prehistoric Human Bone (red. Price, TD) 10–37 (Cambridge University Press. Nowy Jork, 1989).
-
Schoeninger, MJ, DeNiro, MJ i Tauber, H. Stosunek stabilnych izotopów azotu w kolagenie kostnym odzwierciedla morskie i lądowe składniki diety człowieka prehistorycznego. Science 220 , 1381–1383 (1983).
-
Schwarcz, HP, Dupras, T. & Fairgrieve, S. Wzbogacanie 15N w Saharze: w poszukiwaniu globalnego związku. J. Archaeol. Sci. 26 , 629–636 (1999).
-
Szpak, P., Millaire, JF, White, CD i Longstaffe, F. Wpływ nawożenia guanem ptaków morskich i odchodami wielbłądowatych na skład izotopowy azotu w uprawianej w polu kukurydzy ( Zea mays ). J. Archaeol. Sci. 39 , 3721–3740 (2012).
-
Santana-Sagredo, F. i in. Nawóz guano o nazwie „białe złoto” przyczynił się do intensyfikacji rolnictwa na pustyni Atakama od roku 1000 n.e. Nat. Plants 7 , 152–158 (2021).
-
Tykot, R. H., Burger, R. L. i van der Merwe, N. J. Znaczenie kukurydzy w początkowym okresie i wczesnym horyzoncie Peru w dziejach kukurydzy: multidyscyplinarne podejścia do prehistorii, językoznawstwa, biogeografii, udomowienia i ewolucji kukurydzy (red. Staller, JE, Tykot, RH i Benz, BF) 187–197 (Elsevier, Nowy Jork, 2006).
-
Ericson, JE, West, M., Sullivan, C. i Krueger, HW Rozwój uprawy kukurydzy w dolinie Virú w Peru w The Chemistry of Prehistoric Human Bone , (red. Price, TD) 68–104 (Cambridge University Press. Nowy Jork, 1989).
-
Gerdau-Radonić, K., Goude, G., Castro de la Mata, P., André, G., Schutkowski, H. i Makowski, K. Dieta na przedhiszpańskim środkowym wybrzeżu Peru. J.Arch. Nauka. Rep. 4 , 371–386 (2015).
-
Fernandes, R., Millard, AR, Brabec, M., Nadeau, MJ i Grootes, PM Rekonstrukcja żywności przy użyciu sygnałów przenoszonych izotopowo (FRUITS): model bayesowski do rekonstrukcji diety. PLoS ONE 9 (2), e87436 (2014).
-
Fernandes, R., Grootes, PM, Nadeau, MJ i Nehlich, O. Ilościowa rekonstrukcja diety populacji neolitycznej z wykorzystaniem bayesowskiego modelu mieszania (FRUITS): studium przypadku Ostorf (Niemcy). Am. J. Phys. Anthropol. 158 , 325–340 (2015).
-
Colonese, AC i in. Dowody izotopów stabilnych na zróżnicowanie diety w Amazonii przedkolumbijskiej. Sci. Rep. 10 , 16560 (2020).
-
DeNiro, MJ. Pośmiertne zachowanie i modyfikacja stosunków izotopów kolagenu w kościach in vivo w kontekście rekonstrukcji paleodietowej. Nature 317 , 806–809 (1985).
-
Van Klinken, GJ Wskaźniki jakości kolagenu kostnego do pomiarów paleodietologicznych i radiowęglowych. J. Arch. Sci. 26 , 687–695 (1999).
-
Vázquez, VF, Rosales Tham, T., Dillehay TD i Netherly, PJ. Pozostałości fauny. W książce Where the Land Meets the Sea: Fourteen Millennia of Human History at Huaca Prieta, Peru (red. Dillehay, TD) 197–366 (University of Texas, Austin, TX, 2017).
-
Elera , C. Stanowisko Puémape i kultura Cupisnique: studium przypadku początków i rozwoju złożonego społeczeństwa w środkowych Andach w Peru (rozprawa doktorska. Uniwersytet Calgary, Alberta, 1998).
-
Hyland, C., Millaire, J.-F. i Szpak, P. Migracja i kukurydza w dolinie Virú: zrozumienie historii życia poprzez wielotkankową analizę izotopów węgla, azotu, siarki i strontu. Am. J. Phys. Anthropol. 176 , 21–35 (2021).
-
Reitz, EJ Wykorzystanie zasobów w czasie w Paloma Peru. Bull. Fla. Mus. Natl. Hist. 44 , 65–80 (2003).
-
Coutts, KH, Chu, A. i Krigbaum, J. Paleodiet w późnym Peru przedceramicznym: wstępne dane izotopowe z Bandurrii. J. Island Coast. Archaeol. 6 , 196–210 (2011).
-
Sotelo, C. (red.) Cuaderno de Investigación del Archivo Tello nr 9. Paracas WariKayan (Museo de Arqueología y Antropología de la Universidad Nacional Mayor de San Marcos, Lima, 2012).
-
Cadwallader, L. Badanie 1500 lat zmian w diecie w dolinie rzeki Ica w Peru z wykorzystaniem podejścia izotopowego (rozprawa doktorska, Uniwersytet Cambridge, Wielka Brytania, 2013).
-
Finucane, BC Mumie, kukurydza i obornik: wielotkankowa analiza stabilnych izotopów w późnoprehistorycznych szczątkach ludzkich z doliny Ayacucho w Peru. J. Arch. Sci. 34 (12), 2115–2124 (2007).
-
Turner, BL i in. Dieta i zwyczaje żywieniowe na przestrzeni pięciu tysiącleci w regionie Cusco w Peru. J. Arch. Sci. 98 , 137–148 (2018).
-
Kellner, CM i Schoeninger, MJ. Korelacje dietetyczne z rozwojem złożoności społecznej Nasca (1–750 n.e.). Lat. Am. Antiq. 23 , 490–508 (2012).
-
Ikehara, HC, Paipay, JF i Shibata, K. Ucztowanie z Zea Mays na środkowym i późnoformatywnym północnym wybrzeżu Peru. Lat. Am. Antiq. 24 , 217–231 (2013).
-
Goldstein, DJ, Coleman, RC i Williams, PR Jesteś tym, co pijesz. Socjokulturowa rekonstrukcja prehiszpańskiego wykorzystania napojów fermentowanych w Cerro Baúl, Moquegua, Peru w książce Drink, Power, and Society in the Andes (red. Jennings, J. i Bowser, BJ) 133–176 (University Press of Florida. 2009).
-
Gil, AF i in. Dowody izotopowe na kościach ludzkich wskazujące na spadek spożycia kukurydzy w okresie małej epoki lodowcowej w środkowo-zachodniej Argentynie. J. Archaeol. Sci. 49 , 213–227 (2014).
-
Antúnez de Mayolo, S. Ciencia agrícola en el Perú Pre-Colombino w Estudios de la Ciencia en el Perú (red. Yepes, E.) (CONCYTEC, Lima, 1986).
-
Sandweiss, DH, Shady, R., Moseley, M., Keefer, DK i Ortloff, CR Zmiany środowiskowe i rozwój gospodarczy na wybrzeżu Peru w okresie od 5800 do 3600 lat temu. Proc. Natl. Acad. Sci. USA 106 , 1359–1363 (2009).
-
Ortloff, CR i Moseley, ME 2600–1800 p.n.e. Caral: Zmiana środowiska na stanowisku z późnego okresu archaicznego na północno-środkowym wybrzeżu Peru Ñawpa Pacha. J. Andean Archaeol. 32 , 189–206 (2012).
-
Guédron, S. i in. Holoceńskie wahania poziomu wody w jeziorze Titicaca i ich implikacje dla rozwoju społeczno-politycznego w centralnych Andach. Proc. Natl. Acad. Sci. USA 120 , e2215882120 (2023).
-
Robinson, JR Badanie niszy izotopowej: wykorzystanie rKIN do badań izotopów stabilnych w archeologii. J. Archaeol. Method Theory 29 , 831–861 (2022).
-
Koch, PL, Tuross, N. i Fogel, ML Wpływ obróbki próbek i diagenezy na integralność izotopową węglanu w biogenicznym hydroksyapatycie. J. Archaeol. Sci. 24 , 417–429 (1997).
-
Francja, CAM, Sugiyama, N. i Aguayo, E. Ustalanie wskaźnika zachowania minerału bioapatytu kości, zębiny i szkliwa przy użyciu ATR-FTIR. J. Arch Sci. Rep. 33 , 102551 (2020).
-
Tsutaya, T. i Yoneda, M. Rekonstrukcja praktyk karmienia piersią i odstawiania od piersi z wykorzystaniem analizy izotopów stabilnych i pierwiastków śladowych: przegląd. Am. J. Phys. Anthropol. 156 , 2–21 (2015).
-
Marino, BD i McElroy, MB Skład izotopowy atmosferycznego CO2 wnioskowany na podstawie węgla w celulozie roślinnej C4. Nature 349 , 127–131 (1991).
-
DeNiro, MJ i Hastorf, C. A., Zmiana stosunków 15N/14N i 13C/12C w materii roślinnej w początkowych etapach diagenezy: badania z wykorzystaniem okazów archeologicznych z Peru. Geochim. Cosmochim. Acta 49 , 97–115 (1985).
-
Szpak, P. i Chiou, KL Porównanie składu izotopów azotu w zwęglonych i wysuszonych szczątkach botanicznych z północnego Peru. Veg. Hist. Archaeobot. 29 , 527–538 (2019).
-
Fernandes, R. Prosty model (R) do przewidywania źródła węgla w diecie u poszczególnych konsumentów. Archaeometry 58 , 500–512 (2016).
-
Fernandes, R., Nadeau, MJ i Grootes, PM Model oparty na makroskładnikach do analizy transportu węgla z pożywienia w kolagenie kostnym i bioapatycie. Arch. Anthropol. Sci. 4 , 291–301 (2012).
-
Jim, S., Jones, V., Ambrose, SH i Evershed, RP Kwantyfikacja źródeł węgla w makroskładnikach diety dla biosyntezy kolagenu kostnego z wykorzystaniem analizy stabilnych izotopów węgla w naturalnej obfitości. Br. J. Nutr. 95 , 1055–1062 (2006).
-
Pestle, WJ, Crowley, BE i Weirauch, MT Kwantyfikacja zmienności międzylaboratoryjnej w analizie izotopów stabilnych starożytnych szczątków szkieletowych. PLoS ONE 9 (7), e102844 (2014).
Raporty naukowe tom 14 , Numer artykułu: 4582 ( 2024 )
Podziękowanie
Autorzy pragną podziękować Kristie McGrath i Thiago Fossile’owi za recenzję manuskryptu i cenne uwagi dotyczące pierwotnej wersji.
Finansowanie
Projekt ten otrzymał dofinansowanie z programu badań i innowacji Unii Europejskiej „Horyzont Europa” na podstawie umowy o dotację „Maria Skłodowska-Curie” nr 101062179 (Analizy paleodietologiczne pierwszych miast andyjskich: ocena makroskładników w wysokiej rozdzielczości z wykorzystaniem podejścia wieloproksymacyjnego – PACHAMAMA). Prace te zostały również sfinansowane przez projekt TRADITION, będący częścią programu konsolidacyjnego ERC, który otrzymał dofinansowanie od Europejskiej Rady ds. Badań Naukowych (ERBN) w ramach programu badań i innowacji Unii Europejskiej „Horyzont 2020” na podstawie umowy o dotację nr 817911. Prace te stanowią wkład w program „ICTA-UAB María de Maeztu” dla jednostek doskonałości hiszpańskiego Ministerstwa Nauki i Innowacji (CEX2019-000940-M). Niniejsza praca stanowi również wkład w projekt EarlyFoods (Ewolucja i wpływ wczesnych systemów produkcji żywności), który otrzymał dofinansowanie od Agència de Gestió d’Ajuts Universitaris i de Recerca de Catalunya (SGR-Cat-2021, 00527). Fundatorzy nie mieli wpływu na projekt badania, gromadzenie i analizę danych, decyzję o publikacji ani przygotowanie manuskryptu.
Deklaracje etyczne
Konflikty interesów
Autorzy deklarują brak konfliktu interesów.
Informacje dodatkowe
Uwaga wydawcy
Springer Nature zachowuje neutralność w kwestii roszczeń jurysdykcyjnych zawartych w publikowanych mapach oraz powiązań instytucjonalnych.
Prawa i uprawnienia
Otwarty dostęp Niniejszy artykuł jest licencjonowany na podstawie licencji Creative Commons Uznanie autorstwa 4.0 Międzynarodowe, która zezwala na używanie, udostępnianie, adaptację, dystrybucję i reprodukcję na dowolnym nośniku lub w dowolnym formacie, pod warunkiem wskazania autorów i źródła, podania linku do licencji Creative Commons oraz wskazania, czy wprowadzono zmiany. Obrazy lub inne materiały stron trzecich w tym artykule są objęte licencją Creative Commons, o ile nie wskazano inaczej w informacji o autorstwie materiału. Jeśli materiał nie jest objęty licencją Creative Commons, a zamierzony sposób wykorzystania nie jest dozwolony przez przepisy ustawowe lub wykracza poza dozwolony zakres, należy uzyskać zgodę bezpośrednio od właściciela praw autorskich. Aby wyświetlić kopię tej licencji, odwiedź stronę http://creativecommons.org/licenses/by/4.0/ .
Pezo-Lanfranco, L., Colonese, AC Rola rolnictwa i rybołówstwa we wzroście złożoności społecznej w centralnych Andach: perspektywa stabilnych izotopów. Sci Rep 14 , 4582 (2024). https://doi.org/10.1038/s41598-024-55436-4
Link do artykułu: https://www.nature.com/articles/s41598-024-55436-4