


A Okaz nr 3713, kwarcytowe duże narzędzie tnące z warstwy I. B Okaz nr 2486, kwarcytowe duże narzędzie tnące z warstwy III. C Okaz nr 890, bazaltowe narzędzie przypominające tasak z warstwy IV. D Okaz nr 2262, kwarcytowy rdzeń bifacjalny dośrodkowy z warstwy III. E Okaz nr 2318, kwarcyt ząbkowany z warstwy III. F Okaz nr 1197, fonolitowy dwuścienny skrobak boczny z warstwy IV.
Streszczenie
Pytania o to, kiedy wczesni przedstawiciele rodzaju Homo przystosowali się do ekstremalnych środowisk, takich jak pustynie i lasy deszczowe, tradycyjnie skupiały się na Homo sapiens . Tutaj przedstawiamy wielodyscyplinarne dowody z Engaji Nanyori w wąwozie Oldupai w Tanzanii, ujawniające, że Homo erectus rozwijał się w krajobrazach hiperpustynnych milion lat temu. Korzystając z analiz biogeochemicznych, precyzyjnego datowania chronometrycznego, symulacji paleoklimatycznych, modelowania biomów, rekonstrukcji historii pożarów, badań paleobotanicznych, zespołów fauny i dowodów archeologicznych, rekonstruujemy środowisko zdominowane przez półpustynne zarośla. Pomimo tych wyzwań, Homo erectus wielokrotnie zajmował krajobrazy rzeczne, wykorzystując źródła wody i ekologiczne punkty centralne w celu złagodzenia ryzyka. Te odkrycia sugerują, że archaiczni ludzie posiadali ekologiczną elastyczność, którą wcześniej przypisywano wyłącznie późniejszym homininom. Ta zdolność adaptacji prawdopodobnie ułatwiła ekspansję Homo erectusa w suche regiony Afryki i Eurazji, co na nowo określiło jego rolę jako ekologicznego generalisty, który radził sobie w jednych z najtrudniejszych krajobrazów środkowego plejstocenu.
Podobna treść jest oglądana przez innych



Wstęp
Od dawna toczą się dyskusje na temat tego, kiedy rodzaj Homo nabył zdolność adaptacji do życia w ekstremalnych środowiskach, takich jak pustynie i lasy deszczowe 1 , 2 , 3 . Tradycyjnie uważano, że tylko Homo sapiens jest zdolny do długotrwałego zamieszkiwania takich ekosystemów 2 , 3 , a archaiczne homininy były postrzegane jako ograniczone do węższych zakresów 4 , 5 . Jednak dowody biogeochemiczne, paleośrodowiskowe i archeologiczne sugerują, że wczesny Homo miał zdolność adaptacji do zróżnicowanych 6 i niestabilnych środowisk z dna Wielkiego Rowu Wschodnioafrykańskiego 7 i obszarów Afromontane 8 już dwa miliony lat temu. W niniejszym artykule przedstawiamy multidyscyplinarną analizę danych geologicznych, paleontologicznych, archeologicznych, chemicznych i paleobotanicznych z Engaji Nanyori w wąwozie Oldupai w Tanzanii, ujawniając, że Homo erectus przystosował się do zmieniających się hiperpustynnych krajobrazów milion lat temu — na długo przed pojawieniem się naszego gatunku. Ten profil adaptacyjny, charakteryzujący się odpornością w strefach suchych, podważa założenia dotyczące ograniczeń rozprzestrzeniania się wczesnych hominidów i przedstawia H. erectus jako wszechstronnego generalistę i pierwszego hominida, który przekroczył granice środowiskowe w skali globalnej.
Modele systemu ziemskiego 4 , 5 przewidują, że Homo habilis i Homo ergaster (afrykański Homo erectus ) były wysoce wrażliwe na zmienność środowiskową, posiadały ograniczony zasięg ekologiczny i były podatne na cykle precesyjne modulowane ekscentrycznością w paleoklimacie. Tę wrażliwość uznano za dowód, że gatunkom tym brakowało szerokiej zdolności adaptacyjnej potrzebnej do rozwoju w zróżnicowanych ekosystemach, co wzmacnia ich klasyfikację jako specjalistów środowiskowych 4 , 5 , 6 , powszechnie uważanych za zajmujące głównie środowiska mozaikowe obejmujące lasy, tereny zadrzewione i łąki. Gdy te homininy zapuszczały się w regiony o ekstremalnych rocznych opadach — czy to bardzo niskich, czy bardzo wysokich — przetrwanie wymagało wyspecjalizowanych zdolności adaptacyjnych, które nie ujawniły się w pełni aż do pojawienia się Homo sapiens , kiedy to bardziej uogólnione strategie umożliwiły globalny zakres ekologiczny 1 , 2 , 3 . Jednakże ta perspektywa jest uwarunkowana fragmentarycznymi danymi z kluczowych okresów ewolucyjnych, ma ograniczoną rozdzielczość paleoekologiczną na poziomie lokalnym i pochodzi z bardzo małej liczby rozproszonych stanowisk paleoantropologicznych. Ponadto istnieje dodatkowe wyzwanie badawcze w odróżnianiu niszy ekologicznej gatunku hominina — zdefiniowanej jako pełen zakres warunków środowiskowych, które może wykorzystać — od jego elastyczności kulturowej, z jaką reaguje na zmiany środowiskowe. W tym artykule ponownie bada się zdolność adaptacji afrykańskiego Homo erectusa i twierdzi się, że ich nisza ekologiczna nie została ukształtowana przez czynniki abiotyczne, takie jak sam klimat, ale również przez ich zdolność do pokonywania wyzwań fizjologicznych i poznawczych. Integrując te aspekty, kwestionujemy dominujący pogląd na afrykańskiego Homo erectusa jako specjalistę ekologicznego i oferujemy bardziej zniuansowane zrozumienie ich zdolności adaptacyjnych. Ta perspektywa rozszerza naszą koncepcję wczesnych zdolności hominina, wskazując, że rozwój strategii generalistycznych poprzedzał Homo sapiens i nastąpił znacznie wcześniej, niż wcześniej sądzono, zaczynając co najmniej od przejścia środkowoplejstoceńskiego (MPT, 1,2–0,8 Ma).
Aby zbadać, w jaki sposób wahania klimatyczne wpłynęły na zasięg ekologiczny, wzorce rozprzestrzeniania się i technologie wczesnego Homo , przeprowadziliśmy interdyscyplinarne badania w kluczowej lokalizacji wczesnych homininów w strefie równikowej Tanzanii, Engaji Nanyori 9 , 10 , wąwóz Oldupai, podłoże III 11 . Przedstawiamy tutaj duże zestawy danych z nowo wykopanych kontekstów, kompleksową stratygrafię sedymentacyjną i chronologiczne ramy precyzyjnych dat bezwzględnych, które kontekstualizują położenie szczątków Homo erectus znalezionych w latach 60. XX wieku 12 , 13 . Wykorzystując podejście wieloproxy, nasze badania wskazują, że region otaczający wąwóz Oldupai doświadczał warunków półpustynnych obejmujących pierwszą połowę MPT. Roślinność charakterystyczna dla tych półpustyń, w tym taksony takie jak Ephedra spp., obecnie występuje tylko na obszarach położonych tysiące kilometrów na północ, wzdłuż obrzeży Sahary i rozciągających się na Arabię. Dowody te prowadzą nas do wniosku, że wczesne homininy były w stanie przystosować się i przetrwać w środowiskach hiperpustynnych, wykazując szerszą plastyczność ekologiczną niż wcześniej rozumiano. Nasza analiza, obejmująca dane na temat społeczności roślinnych, pożarów lasów, zrujnowanych szczątków fauny i licznych narzędzi kamiennych, ujawnia behawioralną odpowiedź na suchość charakteryzującą się stałą ogniskowością w użytkowaniu ziemi i zaopatrywaniu. W odpowiedzi na dotkliwą suchość podczas MPT, homininy łagodziły ryzyko poprzez strategiczne zamieszkiwanie krajobrazów rzecznych, wielokrotnie zajmując tę samą lokalizację przez tysiąclecia. Ich eksploatacja zasobów fauny i kamieni w pobliżu ujść rzek podkreśla stabilną, adaptacyjną strategię do suchych warunków panujących na wschodnioafrykańskich pustyniach stepowych w tym okresie.
Wyniki
Dynamika rzeczna, stratygrafia sedymentacyjna i chronologia
Podczas osadzania się warstwy III (1,2–0,9 mln lat temu) geografia wschodniego Serengeti uległa znacznym zmianom z powodu procesów tektonicznych i klimatycznych11 , 14 , 15 , 16 , 17. Normalne pękanie uskoków związane z ryftowaniem doprowadziło do zmian w położeniu basenu sedymentacyjnego Oldupai i sieci drenażu rzecznego w jego obrębie11 (rys. 1 , uwaga uzupełniająca 1 ). W rezultacie rzeki przesunęły swoje biegi w kierunku wschodnim i północnym, przecinając słone, wapienne i zasadowe równiny zalewowe11 , 14 , jak pokazano na przykładzie wytrącania się analcymu i kalkretów (rys. uzupełniający 1 ). Jezioro Oldupai doświadczyło epizodów wysychania, na co wskazywały wahania poziomu wody, obecność przerw i geochemiczne markery suszy 15 , 16 , 17 (rys. uzupełniający 2 ). Te fazy wysychania doprowadziły również do powstania warstw pustynnych czerwonych 18 , 19 powstałych w wyniku utleniania osadów zawierających żelazo (rys. 1 , rys. uzupełniające 3 i 4 ).

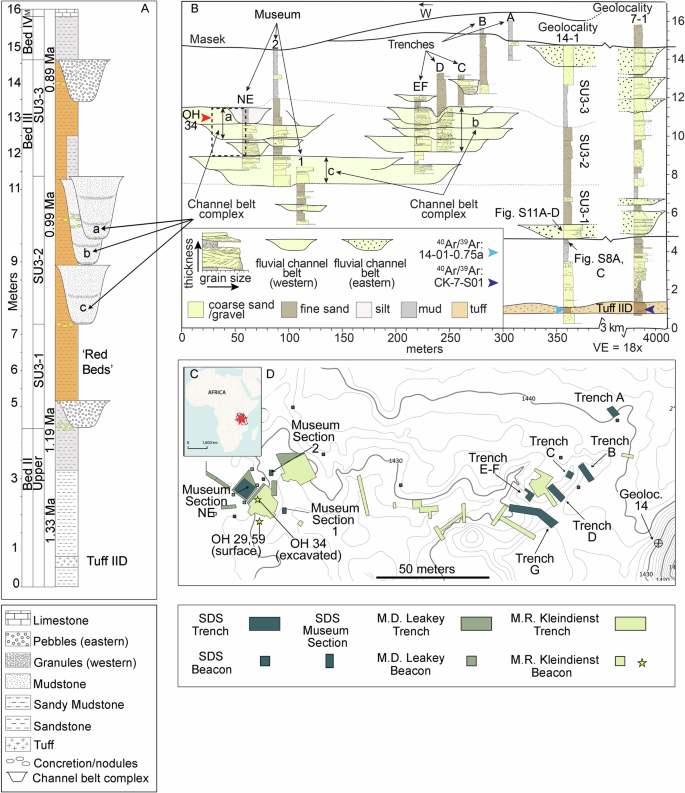
A Diagram stratygraficzny przedstawia litologię, lokalizację kompleksów pasa kanałowego i jednostki stratygraficzne znalezione w warstwach II, III i IV. Ponadto ilustruje on pozycjonowanie stratygraficzne szczątków Homo erectus z lat 60. XX wieku (hominid Olduvai nr 34) 12 , 13 . M Masek. B Przekrój stratygraficzny w Engaji Nanyori rozciągający się od Muzeum Leakeya (zachód) do lokalizacji geologicznej nr 14 (klif Korongo Jumy) i Emurutoto (lokalizacja geologiczna nr 7) na wschodzie. Zakres pionowy tego badania rozciąga się od Tuff IID (warstwa II) do warstw Masek, ze szczególnym uwzględnieniem osadów pasa kanałowego z warstwy III. Przybliżona lokalizacja pionowa hominidu Olduvai 34 jest oznaczona czerwoną strzałką. SU jednostka stratygraficzna. C , D Lokalizacja w Afryce Wschodniej oraz mapa stanowiska pokazująca położenie wykopalisk archeologicznych wykonanych przez Kleindiensta w latach 1962–1970 , Leakeya w latach 1970–1970 i nasz zespół w 2022 r.
Obraz w pełnym rozmiarze
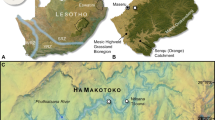
Zbieg rzek utworzył się na północ od Engaji Nanyori podczas MPT, oznaczając jedno z niezwykle rzadkich stanowisk artefaktów/skamieniałości z tego etapu. Lokalizacja typu dla Bed III 9 , 10 zawiera dowody na istnienie dwóch łączących się działów wodnych o bardzo różnych biomach: wyżyn wulkanicznych na południowym wschodzie i równin Serengeti na zachodzie (Rys. 2 , Ryc. uzupełniający 5 ). Osady pasa kanałowego są różnicowane na dwa typy według pochodzenia, orientacji i rozmieszczenia osadów (Rys. uzupełniający 6 ). Stałe i efemeryczne strumienie pozostawiły złożony zapis sedymentologiczny zmieniających się środowisk, wyrażony w pakietach sześciu facji (Rys. 1 , Notatka uzupełniająca 1 , Rys. uzupełniający 2 , Rys. 3 ): konglomerat wulkanoklastyczny (facja 1), pstrokaty czerwony mułowiec (facja 2), czerwonobrązowy mułowiec (facja 3), drobnoziarnisty do gruboziarnistego piaskowca (facja 4), jasnoszary do różowego mułowca (facja 5) i szary mułowiec (facja 6). Facje te reprezentują meandrujące kanały rzeczne o głębokości ~1 m/szerokości ~20 m i towarzyszące im równiny zalewowe, które są wyrażone w trzech jednostkach stratygraficznych: 3–1, 3–2 i 3–3 (Rys. 1 ). W obrębie jednostki 3–2 niedawno zidentyfikowano osady trzech odrębnych pasów kanałowych: A, B i C (rys. 1 , uwaga uzupełniająca 1 ), charakteryzujące się piaszczystymi do żwirowych łach punktowych i drobnoziarnistymi osadami opuszczonych kanałów. Najstarszym jest C (odpowiednik „niższego grubego szarego piasku” Leakeya 10 ), składający się z osadów pasa kanałowego połączonych pionowo i bocznie. Osady pasa kanałowego B mają 125 m szerokości i charakteryzują się żwirami zabarwionymi żelazem („drobnoziarnisty piasek żelazisty” Leakeya 10 ). Pas kanału A jest najmłodszy i dominują w nim żwirowe piaskowce warstwowane krzyżowo (wykopy Kleindiensta A i B 9 ), które ku górze są cienkie – te drobniejsze osady dostarczyły skamieniałości Homo erectus (hominid Olduvai, OH34) 12 , 0,5 m poniżej mułowców Leakeya z dołkami i bruzdami (rys. uzupełniający 4B ) 10 i około 5 m na południowy wschód od obecnego obszaru badawczego (rys. 1 , rys. uzupełniający 7 ).

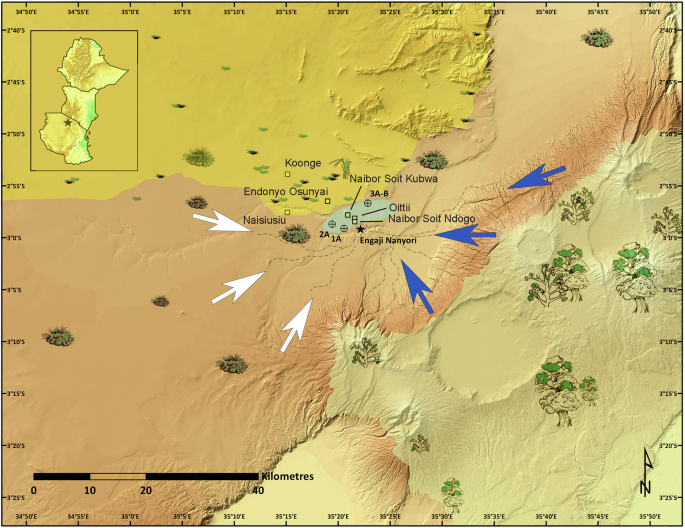
Dominującą grupą roślin w wąwozie Oldupai w początkowej fazie przejścia środkowoplejstoceńskiego były nagonasienne. Wulkaniczne wyżyny (prawy dolny róg, jasnozielone cieniowanie) były domem dla drzew iglastych, podczas gdy wschodnia część Serengeti była pokryta krzewami efedroidów, roślinami jednoliściennymi i trawami C 4. Engaji Nanyori (symbol gwiazdy) znajdowało się u zbiegu wschodnich (niebieskie strzałki) i zachodnich (białe strzałki) strumieni. Miejsca wierceń 16 są pokazane jako symbol celu. Źródła kwarcytu wykorzystywane przez homininów acheulskich w Engaji Nanyori są oznaczone jako kwadraty.
Obraz w pełnym rozmiarze
Ryc. 3: Zdjęcia odsłonięć i powierzchni wykopów przedstawiających najważniejsze formacje geologiczne w Engagi Nanyori.


Tuf IID odsłonięty na klifie JK (rys. uzupełniający 8 ). B Wulkaniczny konglomerat klastowy osadów dolnego pasa korytowego w latach 14–1. C Margines osadu pasa korytowego w wykopie muzealnym nr 1-NE wskazujący na głębokość kanału ~1 m. D Wapienne ryzolity w czerwonawym mułowcu typowym dla osadów zalewowych warstwy III. E Granulowany piaskowiec typowy dla zachodniego źródła osadów pochodzących z podłoża (patrz rys. uzupełniający 6E, F ). F Krzyżowo warstwowane piaskowce pasa korytowego 2 z wykopu D. Kierunki paleoprądów wyprowadzono z kierunku nachylenia zalewisk. G Bezstrukturalny biały do różowawego mułowiec pasa korytowego A w wykopie muzealnym nr 1-NE.
W miejscu badań, Bed III zgromadziło 10 m osadu w ciągu 300 000 lat 20 (Rys. 1A ). Osady Bed III są łatwo rozpoznawalne po ich wyraźnym ceglastoczerwonym kolorze, znajdującym się 3,5 m nad Tuff IID (górne Bed II; Rys. 1A, B , Rysunki uzupełniające 2 i 8 ). Sprzeczne szacunki wieku dla kluczowego znacznika Tuff IID 21 zostały rozwiązane dzięki nowemu wiekowi 40 Ar/ 39 Ar wynoszącemu 1,332 ± 0,020 Ma (2σ) z pobliskiego odsłonięcia we wschodniej paleobazinie, który jest fizycznie i geochemicznie skorelowany zarówno z Engaji Nanyori (rys. 4, rysunki uzupełniające 8–10 ) , jak i zachodnim wąwozem 22 (patrz Metody, Tabela uzupełniająca 1 , https://doi.org/10.20383/103.0901 ). Nowy wiek radiometryczny i korelacja ustanawiają spójny wiek w całym basenie dla Tuff IID. Korzystając z naszego nowego wieku z Tuff IID jako kotwicy chronostratygraficznej, ograniczamy poprzednie modele 20 i sugerujemy wiek 1,19 ± 0,06/0,07 Ma (1σ) dla podstawy Bed III (rys. 4 ). Ta absolutna data jest niezbędna pośród bardzo skąpych dat dostępnych dla warstwy III. Nie tylko wyjaśnia ona wcześniejsze niepewności i rozbieżności21 , ale także rozszerza chronologię najniższej warstwy III do początku MPT, z poprzedniego szacunku 1,14 ± 0,07 Ma20 , umieszczając jednocześnie Homo erectus12 , po raz pierwszy, w środku warstwy III (jednostka stratygraficzna 3-2) na 0,99 ± 0,06/0,07 Ma (1σ) (rys. 1B i 4 ). Ta poprawiona ocena wieku hominina OH34 opiera się na jego pozycji stratygraficznej, określonej w kolumnie geologicznej i popartej modelowaniem głębokości wieku.

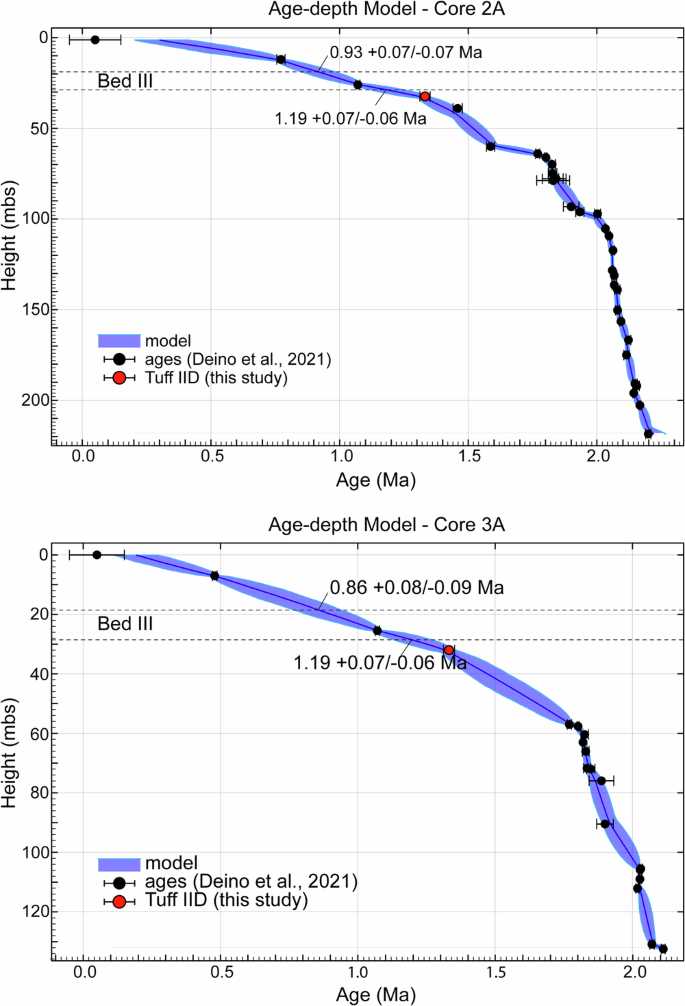
Nowa data Tuff IID (czerwone kółko) jest naniesiona 3,5 m poniżej warstwy III jako odpowiedni horyzont w rdzeniach 2 A i 3A 20 . Wykorzystując wcześniej opublikowane dane wiekowe 20 i kod modelu bayesowskiego wieku-głębokości Chron.jl, włączamy nowy wiek Tuff IID do modelowania relacji wiek-głębokość w obu rdzeniach. Model szacuje wiek podstawy i wierzchołka warstwy III przy 95% CI. Średni wiek podstawy warstwy III z obu rdzeni wynosi 1,19 ± 0,07/0,06 Ma, różniąc się od poprzednich modeli, które szacowały go na 1,14 Ma 20 . Wiek wierzchołka warstwy III jest szacowany na 0,93 dla rdzenia 2 A i 0,86 ± 0,08/0,09 Ma dla rdzenia 3 A, zgodnie z poprzednimi modelami 20 . Wiek skamieniałości Homo erectusa , na podstawie miejsca, jakie ustaliliśmy dla nich w kolumnie geologicznej, szacuje się na 0,99 ± 0,07/0,05 Ma.
Degradacja ekologiczna w okresie przejścia środkowoplejstoceńskiego
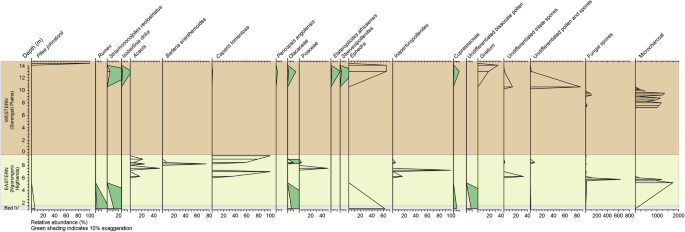
W regionach, w których rozległa ewapotranspiracja w zamkniętych basenach ogranicza dostępność słodkiej wody, często pojawiają się jałowe ziemie, co prowadzi do powstawania pustyń stepowych lub środowisk półpustynnych23 , 24 , 25. W kontekście Engaji Nanyori dowody na przystosowanie się Homo erectus do ekstremalnych i bardzo zmiennych klimatów środkowoplejstocenu stają się widoczne pośród suchych warunków, zmniejszonej pokrywy roślinnej i częstych zaburzeń ekologicznych spowodowanych pożarem. Analiza pyłków kopalnych z półpustynnej społeczności roślinnej charakteryzującej się znacznym udziałem nagonasiennych w porównaniu do okrytonasiennych (10:1), przy czym dominującym rzędem są Gnetales (ryc. 5 ) , ujawnia obecność co najmniej sześciu gatunków krzewów efedroidów suchych (tabela uzupełniająca 2 , ryc. 6 , uwaga uzupełniająca 2 ). Co godne uwagi, w zapisie pyłkowym przeważają pyłki Ephedripites (ryc. 5E, F ), wytwarzane przede wszystkim przez Ephedra , rodzaj znany z odporności na suszę, charakterystycznej dla ekosystemów perysaharyjskich 26 , 27 , 28 .
Ryc. 5: Zasięg geograficzny biomów Ephedraceae i Saharo-Sindian w niedawnych czasach29 i ich hipotetyczny zasięg podczas przejścia środkowoplejstoceńskiego w Afryce. Kluczowe typy pyłków.

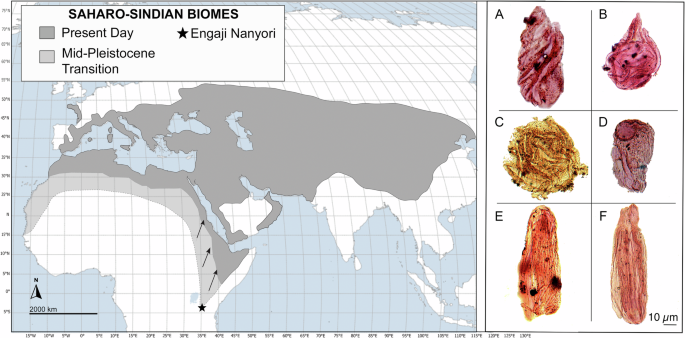
Symbol gwiazdki pokazuje lokalizację Engaji Nanyori. Mapa bazowa utworzona przez Rennę Truong, Spatial and Numeric Data Services w Libraries and Cultural Resources na University of Calgary, 22 stycznia 2024 r., przy użyciu technologii ESRI. Wybrany pyłek: A Gnetaceaepollenites clathratus (Próbka: Engaji Nanyori 2017, wykop B: IIIA; preparat R-3986-15/OTC, Anglia, współrzędne wyszukiwarki: T22/2). B Elateroplicites spp . (Próbka: Engaji Nanyori 2017, wykop B: IB; preparat R-3986-14/OTC, Anglia, współrzędne wyszukiwarki: S40/1). C Gnetaceaepollenites diversus (Nazwa próbki: Engaji Nanyori 2017, wykop B: IB; preparat R-3986-14/OTC, Anglia Współrzędne wyszukiwarki: R8/2). D Steevesipollenites multilineatus (Nazwa próbki: Engaji Nanyori 2017, wykop B: IB; preparat R-3986-14/OTC, Anglia Współrzędne wyszukiwarki: L18/4). E Ephedra spp. (Nazwa próbki: Engaji Nanyori 2017, wykop A: IV; preparat R-3986-12/OTC; Anglia Współrzędne wyszukiwarki: L23/4). F Ephedra spp. (Nazwa próbki: Engaji Nanyori 2017, wykop B: IIIa; preparat R-3986-15/OTC, Anglia Współrzędne wyszukiwarki: M18/1).

Osady pochodzące ze wschodu i zachodu.
Według naszej wiedzy, odkrycie to oznacza pierwsze i najbardziej wysunięte na południe wystąpienie Ephedra poza strefą saharyjsko-arabską, co wskazuje, że w odpowiedzi na zwiększoną suchość podczas MPT, wąwóz Oldupai, na 3°S, został otoczony roślinnością saharyjsko-sydyńską, rozciągającą się na ponad 1000 km na południe od obecnej granicy 3°N 27 , 28 , 29 , 30 (Rys. 5 ). Aby dalej badać paleośrodowisko Wschodnioafrykańskiego Ryftu podczas MPT i ocenić rozpowszechnienie warunków hiperpustynnych, zastosowaliśmy te same ramy modelowania paleoklimatyczno-roślinnego, co w niedawnych badaniach systemu Ziemi 4 , 5 . Podczas gdy modele o niższej rozdzielczości mogą sugerować, że indeks przydatności siedlisk homininów przekroczył 0,5 4 , 5 , nasza oparta na badaniach terenowych synteza danych paleośrodowiskowych dostarcza licznych dowodów na wysoką suchość, w tym oznaki wysychania jezior, gleby słone/zasadowe, formowanie się pustynnych czerwonych złóż i wzmożoną aktywność pożarów w wąwozie Oldupai. Ponadto, wykorzystując ich dane paleoklimatyczne z symulacji pCESM-3Ma i modelu BIOME4, wygenerowaliśmy projekcje regionalnego biomu o wysokiej rozdzielczości (1 km) (Rys. 7 , Ryc. uzupełniający 11 ). Wyniki wskazują, że MPT charakteryzował się naprzemiennymi fazami mezofitycznych i kserofitycznych społeczności roślinnych, przy czym najbardziej suche warunki występowały podczas MIS 30, w szczególności około 1,043 miliona lat temu. Symulacje te ujawniają znaczącą ekspansję ekosystemów półpustynnych w Afryce równikowej podczas MPT, szczególnie w obszarze między wąwozem Oldupai a południowo-wschodnią częścią Jeziora Wiktorii, gdzie półpustynie zastąpiły obecne lasy i łąki. Ponadto cała kotlina Turkana jest przedstawiana jako prawdziwa pustynia, pozbawiona roślinności i wód powierzchniowych.

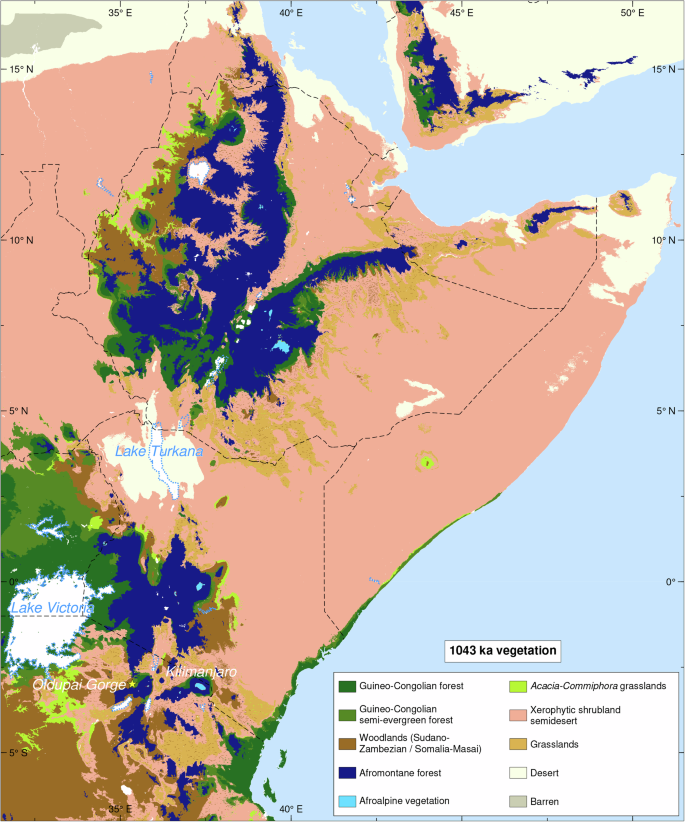
Biomy symuluje się przy użyciu globalnego modelu roślinności BIOME4 opracowanego na podstawie symulacji paleoklimatycznej pCESM.3 Ma.
Analiza stabilnych izotopów węgla w szkliwie zębów roślinożerców wskazuje na dietę zdominowaną przez trawy C4 ( Tabela uzupełniająca 3 ), które są przystosowane do wysokich temperatur napromieniowania (Rys. uzupełniający 12 ), co kontrastuje ze mieszaną ścieżką fotosyntezy C3 – C4 obserwowaną w basenie Oldupai na początku plejstocenu31 . Ponadto badanie gleb zalewowych z facji 2 pokazuje fitolity z krzewów drzewiastych i roślin jednoliściennych (Rys. uzupełniające 13 i 14 ). Zostały one osadzone w warunkach dużego parowania i wytrącania węglanów, co doprowadziło do nukleacji zeolitów32 na fitolity i fosylizacji korzeni palowych (Rys. uzupełniający 15 ), przypominających te z gatunków Ephedra28 . Ponadto obecność średniołańcuchowych normalnych alkanów ( n – C23 ) (rys. uzupełniający 16 ) sugeruje istnienie wodnych roślin zielnych w pobliżu aktywnych, meandrujących kanałów.
Zapis osadowy z Engaji Nanyori dostarcza opisu dawnych pożarów lasów, rozpoznawalnych za pomocą markerów molekularnych i mikroskopowych. Analiza węglowodorów nasyconych ujawnia rozgałęzione i cykliczne alkany z wyraźnym nieparzystym-parzystym wzorem węgla (rys. 8 , uwaga uzupełniająca 3 , rysunki uzupełniające 16 i 17 , tabela uzupełniająca 4 ). Nadzwyczajna przewaga homologów o krótkich łańcuchach i parzystych numerach węgla w rozkładach n-alkanów jest oznaką niepełnego spalania biomasy, co jest wspólną cechą pozostałości po pożarach lasów33 . Podczas zwęglania długołańcuchowe n-alkany ulegają znacznej redukcji, co prowadzi do krótkołańcuchowych n-alkanów o parzystych numerach węgla (m/z 85), zwykle przy C 18 lub C 16 . Współwystępowanie rozgałęzionych i cyklicznych alkanów (dominujące piki w m/z odpowiednio 127, 141/m/z 68, 82) wykazujących wyraźną przewagę węgla nieparzystego/parzystego dodatkowo potwierdza pogląd, że procesy indukowane ciepłem przyczyniają się do obserwowanych wzorców n-alkanów (rys. uzupełniający 17 ). Innym dowodem wzmacniającym pochodzenie pirogeniczne jest stosunek niepodstawionych wielopierścieniowych węglowodorów aromatycznych (WWA) do ich alkilowanych odpowiedników (P/∑MP). Na przykład rozpowszechnienie fenantrenu, głównego składnika w tworzeniu WWA, w porównaniu do alkilowanych metylofenantrenów, pomaga ustalić obecność WWA pochodzących ze spalania w całym miejscu, przy czym średnie stosunki wahają się od 1,03 do 1,43, co potwierdza zdarzenia spalania. Ponadto stosunek związków fluorantenu (Fl) do pirenu (Py), ze średnią 0,67, jest sygnaturą pirogenicznych WWA, co stanowi dowód na występowanie pożarów w tym rejonie w przeszłości. Na koniec, aktywność pożarów na skalę regionalną potwierdza obfitość osadowego mikroskopijnego węgla drzewnego, z wahaniami reżimów pożarowych o różnej intensywności (Rys. uzupełniający 18 , Tabela uzupełniająca 5 ).
Rys. 8: Nałożone na siebie chromatogramy jonów masy względem ładunku (m/z) 178, 192 i 202, ilustrujące rozkład fenantrenu, metylofenantrenów, fluorantenu i pirenu w próbkach z Engaji Nanyori East i West.

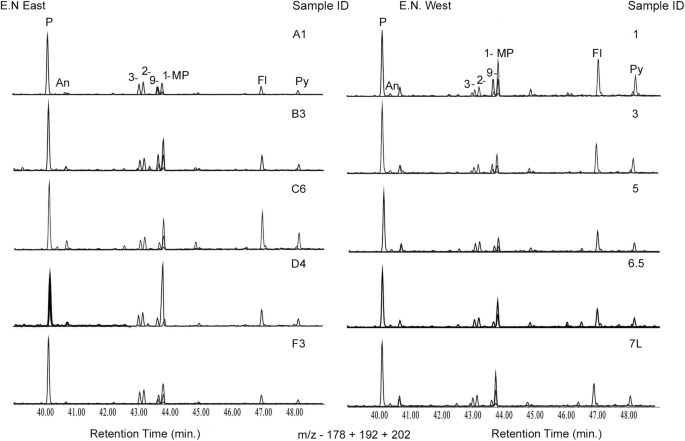
P fenantren, MP metylofenantreny, Fl fluoranten, Py piren. Antracen. Oś pionowa przedstawia względną obfitość w stosunku do ilości próbki wstrzykniętej do instrumentu. Aby obliczyć stężenia, używamy wewnętrznego standardu.
Reakcje na suszę
W hiperpustynnym basenie Bed III aktywność hominidów koncentrowała się głównie na miejscu, gdzie świeża, płynąca woda byłaby częstsza ze względu na obecność dwóch systemów rzecznych o radykalnie różnych dorzeczach. Opady w obu dorzeczach zapewniałyby odpływ rzeki i zwiększały prawdopodobieństwo dostępności wody w przypadku jednego systemu drenażowego. Strategiczny wybór siedliska wokół punktów ogniskowych rzeki zgromadził dowody długiej historii produkcji narzędzi kamiennych do przetwarzania tusz zwierzęcych. W latach 60. i 70. Engaji Nanyori dostarczył bogactwo przerobionych artefaktów i skamieniałości wydobytych z kontekstów wysokoenergetycznych 9 , 10 , w tym kanałów, łach punktowych i podwodnych wydm, typowych dla facji 3. Poprzednie prace interpretowały te pozostałości jako dowód na to, że hominidy obozowały nad płynącą wodą. (Zrekonstruowaliśmy wymiary czynnego kanału jako mierzące 1 m głębokości i 20 m szerokości.) Jednakże pokazujemy tutaj, że pierwotnym siedliskiem preferowanym przez hominidy i głównym źródłem wszystkich materiałów kulturowych były kręte środowiska przypominające stawy facji 5, rozwinięte bocznie do strumienia przez opuszczone koryta rzeczne po ich oderwaniu. Były one powszechne po zachodniej stronie Engaji Nanyori, gdzie znajduje się najbogatsza akumulacja archeologiczna i które służyły jako punkty centralne zarówno dla poprzednich (Kleindienst 9 , Leakey 10 ) i nowych prac. Te preferowane siedliska stawowe były kiedyś szeroko rozpowszechnione, ale zostały zachowane tylko wyjątkowo ze względu na ciągłą przebudowę i erozję spowodowaną migracją nowych kanałów przez krajobraz. W przeciwieństwie do wtórnych, wysokoenergetycznych kontekstów zgłoszonych przez Kleindienst 9 i Leakey 10 , mulisty mułowiec z opuszczonych kanałów, które przedstawiamy, jest niskoenergetyczny. To środowisko sedymentacyjne (rys. uzupełniające 3 i 4B ) wypełnione było drobnoziarnistymi osadami z późniejszego przepływu ponad brzegiem podczas zdarzeń wysokiego przepływu, grzebiąc setki narzędzi i kości zwierzęcych w pierwotnej pozycji i doskonałych warunkach, bez preferencyjnej orientacji kamieni lub skamieniałości (rys. uzupełniający 19 ). Masywne do słabo warstwowanych i dobrze posortowane osady zachowały wiele paleopowierzchni, w tym „dołki i bruzdy” (rys. uzupełniający 4B ), dla których Leakey zbudował muzeum w latach 70. 10 . Skomplikowane zagłębienia, bruzdy i rynny (rys. uzupełniający 4B ) były małymi dołkami i śladami trakcji ściśle przypominającymi te znalezione w piaskowcach Grupy Kilwa 34 , przypisywanymi codziennym ruchom bezkręgowców po mokrym mule zachowanym jako śladowe skamieniałości.
Homo erectus przyniósł duże ilości płyt kwarcytowych do opuszczonych kanałów i brzegów stawów facji 5, jednocześnie pozyskując fonolit i bazalt z otoczaków koryta rzeki (rys. 3B ). Geochemiczne odciski palców pokazują, że większość dostarczanych skał do produkcji narzędzi była ukierunkowana na odsłonięcia metamorficzne ograniczone do korytarza oddalonego o dwa do trzynastu kilometrów stycznie wzdłuż ujścia rzeki (rys. 2 , rysunki uzupełniające 20–23 , uwaga uzupełniająca 4 ). Wyróżniono cztery warstwy (I–IV) na podstawie kryteriów sedymentologicznych, stratygraficznych i archeologicznych ( rysunki uzupełniające 24–29 ) . Zestawy narzędzi kamiennych Engaji Nanyori pochodzą z kompleksu acheulskiego (ryc. 9 ), charakteryzującego się gęstymi skupiskami9 , 10 ( dziedzictwo + zgłaszane w niniejszym dokumencie ≥22 000 sztuk na powierzchni >300 m2 ) i po raz pierwszy łączącego lokalne trajektorie kulturowe wczesnoplejstoceńskich homininów w wąwozie Oldupai, takich jak Homo habilis i Paranthropus boisei (warstwa I–II, 2,0–1,3 mln lat), z gatunkami takimi jak Homo erectus ze środkowego plejstocenu (warstwa III–IV: 1,2–0,9 mln lat) 10 . Facja 5 rejestruje kompletne sekwencje kruszenia poprzez mocne uderzenia, o czym świadczą odłupki korowe, kamienie młoteczkowe i rozproszone cebulki (uwaga uzupełniająca 5 ), pokazujące redukcję kamieni na miejscu. Spośród sześciu metod eksploatacji kamienia (Tabela uzupełniająca 6–8 ) dominowała metoda bifacjalna ortogonalna, unipolarna podłużna i bifacjalna dośrodkowa.

Analiza głównych składowych wewnątrzregionalnych (PCA) stanowisk reprezentujących różne siedliska homininów, strategie utrzymania i kluczowe markery technologiczne (Rys. uzupełniający 30 , Tabele uzupełniające 9–17 ) pokazuje ciągłość między Engaji Nanyori a wcześniejszymi przemysłami oldowańskimi i acheulskimi. Biorąc pod uwagę całą sekwencję obróbki kamienia, PCA wykazuje ścisłe dopasowanie techniczne do tych stanowisk (PC1-2 = 98,62% wariancji), szczególnie w strategiach redukcji rdzenia (PC1-2 = 83,70% wariancji), odzwierciedlając wspólne ramy technologiczne. Jednakże Engaji Nanyori wykazuje wyraźną rozbieżność w typach narzędzi, charakteryzującą się wyższym udziałem narzędzi retuszowanych (rylców, ząbków i skrobaków: PC1-2 = 82,43% wariancji). Ta zwiększona reprezentacja narzędzi retuszowanych, prawdopodobnie używanych do zadań związanych z rzeźnictwem, sugeruje, że Engaji Nanyori funkcjonowało jako wyspecjalizowane miejsce przetwarzania i eksploatacji zasobów. Ten wzór zachowania wskazuje, że podczas gdy Homo erectus utrzymywał spójne strategie łupania w różnych miejscach, rozwinął wyspecjalizowane narzędzia w celu optymalizacji działań związanych z utrzymaniem się w niszach suchych lądów.
Obfitość kości zwierzęcych gromadzonych antropogenicznie sugeruje udane, powtarzalne wykorzystanie stanowiska Engaji Nanyori przez Homo erectus w celu pierwotnego i natychmiastowego dostępu do części osiowych i kończynowych dorosłych osobników wołowatych o wadze <300 kg. Duże ilości kości zwierzęcych (dziedziczne + niniejszy artykuł ≥43 000 okazów na powierzchni >300 m2 ) 35 , 36 , 37 również wskazują na trwałą historię użytkowania stanowiska. Trójwymiarowe współrzędne wykopanych skamieniałości i kości są dodatnio skorelowane między nimi (Tabela uzupełniająca 18 , Rys. uzupełniający 28 , Uwaga uzupełniająca 6 ). To, że ludzie byli głównym drapieżnikiem i zbieraczem szczątków zwierzęcych, niezależnie wskazują niskie poziomy zniszczenia kości mięsożerców i skąpe ślady zębów. Mięso było intensywnie rozbierane (Rys. 10 ), Rys. uzupełniające. 31 – 37 ), z kośćmi podtrzymującymi liczne ślady nacięć wskazujące na obdzieranie, obdzieranie ze skóry i rozczłonkowanie. Ekstrakcja szpiku kostnego jest wnioskowana z antropogenicznych dołków i nacięć pouderzeniowych, związanych ze świeżymi złamaniami. Ponad połowa skamieniałości w niniejszym badaniu była taksonomicznie identyfikowalna i ujawniła zespół o niskiej różnorodności, w którym dominują bovidy (Uwaga uzupełniająca 7 , Tabele uzupełniające 19 – 23 ). Pozostałości faunistyczne obejmują taksony wodne (Crocodylidae, Hippopotamidae) i taksony siedlisk otwartych (Reduncini, Antilopini, Equidae). Taksony zależne od wody są migrującymi lub odpornymi mieszkańcami stawów w okresach suszy, takimi jak żółwie i sumy.


A Skośne, równoległe ślady nacięć na przedniej powierzchni korowej kości długiej; ID okazu: 192. EN Museum Trench nr 1, warstwa IV. B Skośny ślad nacięcia na fragmencie kości długiej; ID okazu 321. EN Museum Trench nr 1, warstwa IV. C Skośne, równoległe ślady nacięcia na fragmencie kości długiej; ID okazu 881. EN Museum Trench nr 1, warstwa IV. D Trójwymiarowy profil skośnych i podłużnych śladów nacięć na fragmencie kości długiej, ID okazu 952. EN Museum Trench nr 1, warstwa IV. E Skośny ślad nacięcia na fragmencie kości długiej; ID okazu 191. EN Museum Trench nr 1, warstwa IV. F Poprzeczny ślad nacięcia na fragmencie kości długiej; ID okazu 54. EN Museum Trench nr 1, warstwa IV.
Dyskusja
Rozszerzanie zasięgu i rozprzestrzenianie się w strefie Saharo-Sindian
Symulacje modeli paleoklimatycznych i roślinności 4 , 5 sugerują, że warunki środowiskowe w północnej Tanzanii mogły być umiarkowanie sprzyjające zasiedleniu przez homininy w okresie MIS 30. Wieloproksymacyjne, oparte na terenie rekonstrukcje paleośrodowiskowe dla regionu Oldupai, które tutaj przedstawiamy, nie potwierdzają tych symulacji, a zamiast tego sugerują, że warunki charakteryzowały się ekstremalną suchością. Modele biomów 4 , 5 przeceniają zasięg terenów leśnych, ponieważ znajdujemy niewiele dowodów na tę roślinność w analizowanych przez nas sekwencjach biogeochemicznych. Ponadto w okresie MIS 30 wąwóz Oldupai charakteryzuje się wysychaniem jezior, glebami słonymi/zasadowymi, formowaniem się pustynnych czerwonych złóż i zwiększoną liczbą pożarów lasów. W takich warunkach środowiskowych krajobraz był zdominowany przez kserofityczne zarośla efedroidów i nie byłby ekologicznie odpowiedni dla terenów leśnych. Pomimo wyzwań stawianych przez tereny suche, prawdopodobnie odegrały one kluczową rolę w warunkowaniu wczesnych ludzkich zachowań, technologii, rozproszenia i ekspansji ekologicznej w Afryce, Lewancie i Arabii. Nowe zestawy danych z Engaji Nanyori potwierdzają wniosek z badania, że Homo erectus wykazywał znacznie większą zdolność adaptacji do różnych ekologii niż wcześniej sądzono, ze zdolnością do trwałego zajmowania ekstremalnych środowisk. Homininy wykazały się wnikliwym i empirycznym zrozumieniem otaczającej ich ekologii poprzez połączenie adaptacyjnych strategii żerowania, używania narzędzi, eksploatacji środowiska i praktyk zarządzania zasobami dostosowanych do ich specyficznego kontekstu ekologicznego. Engaji Nanyori jest najwcześniejszym przykładem afrykańskiego Homo erectus dostosowanego do narastającej suchości i dużej zmienności klimatycznej początkowego okresu MPT. Na podstawie skali czasu sedymentacji pasów trzech kanałów obejmujących pozostałości paleoantropologiczne i archeologiczne, obejmujących od 1000 do 10 000 lat każdy 38, możemy wnioskować o przedłużonej cykliczności ludzkiej okupacji pustyń stepowych północnej Tanzanii. Regularne użytkowanie ekosystemów podatnych na nawracające zakłócenia sugeruje długotrwałą strategię elastycznego wyboru siedliska. Ta strategia prawdopodobnie czerpie swoją adaptacyjną przewagę z reorganizacji wykorzystania zasobów wywołanej degradacją środowiska i fragmentacją. Priorytetem jest zakładanie obozów w pobliżu ujścia rzek i podkreśla znaczenie stawów dla dostępu do zasobów wodnych w trudnych warunkach, zamiast polegać wyłącznie na zmianach w technologii kamiennej. Acheulean Engaji Nanyori przedstawia najwcześniejsze zajęcie strefy ekoklimatycznej Saharo-Sindian, jednego z największych i najbardziej zróżnicowanych regionów łączących Afrykę Wschodnią z Afryką Północną i Zachodnią, wraz z Półwyspem Arabskim. Nabyta zdolność do zamieszkiwania w siedliskach marginalnych stanowiła podstawę niezwykłej zdolności adaptacyjnej, która charakteryzowała Homo podczas MPT i prawdopodobnie umożliwiła szerokie rozproszenie Homo erectus w nowych, zróżnicowanych i ekstremalnych ekosystemach w całej Eurazji. Engaji Nanyori wskazuje na zmianę paradygmatu w zakresie rozumienia globalnego osadnictwa i rozmieszczenia ekologicznego Homo erectus.
Metody
Charakterystyka geochemiczna próbek geologicznych
Zobacz https://doi.org/10.20383/103.0901, tabelę uzupełniającą 1 (warunki analizy za pomocą mikrosondy elektronowej na karcie konfiguracji).
40 Ar/ 39 Ar
Wszystkie prace analityczne wykonano na Uniwersytecie Manitoby przy użyciu wielokolektorowego spektrometru masowego Thermo Fisher Scientific ARGUSVI, połączonego ze stalową linią ekstrakcyjno-oczyszczającą Thermo Fisher Scientific i laserem CO2 Photon Machines (55 W) Fusions 10.6. Izotopy argonu mierzono przy użyciu następującej konfiguracji: 40 Ar (H1; rezystor 1 × 1013 Ω), 39 Ar (AX; rezystor 1 × 1013 Ω), 38 Ar (L1; rezystor 1 × 1013 Ω), 37 Ar (L2; rezystor 1 × 1013 Ω) i 36 Ar (kompaktowa dyskretna dynoda [CDD]). Czułość pomiarów argonu wynosi ~6,3 × 1017 moli/fA, jak określono na podstawie zmierzonych alikwotów Fish Canyon Sanidine 39 , 40 . Standardy i niewiadome umieszczono w 2 mm głębokich dołkach w 18 mm średnicy aluminiowych dyskach, przy czym standardy umieszczono strategicznie, tak aby można było ocenić boczne gradienty strumienia neutronów na dysku. Regresje planarne dopasowano do danych standardowych, a parametr fluencji neutronów 40 Ar/ 39 Ar, J, interpolowano dla niewiadomych. Wszystkie próbki napromieniowano w wyłożonym kadmem, rdzeniowym urządzeniu CLICIT reaktora Oregon State University TRIGA. Czas napromieniowania wynosił 3 godziny, a wykorzystano sanidynę Fish Canyon (28,201 Ma) 40 . Astronomicznie skalibrowany sanidyn ryolitowy Alder Creek (1,18342 ± 0,00069 Ma) 41 został użyty jako standard wtórny. Osiem z dziesięciu analiz (Tabela S1 ) daje wiek 1,181 ± 0,002 Ma (2 õ; MSWD = 3,52) w stosunku do sanidyny Fish Canyon. Napromieniowane próbki umieszczono w tacy na próbki Cu z nakrywką KBr w linii ekstrakcyjnej o wysokiej próżni ze stali nierdzewnej i wypalano lampą podczerwoną przez 24 godziny. Pojedyncze kryształy albo zespolono za pomocą lasera przez jedną minutę, a gazy reaktywne usunięto po ~1 minucie za pomocą kriochłodziarki JANIS w ~−120 °C, pochłaniacza NP-10 SAES w temperaturze pokojowej i pochłaniacza GP50 (stop C50 ST101) w 450 °C przed wpuszczeniem do spektrometru masowego ARGUSVI przez rozprężanie. Pięć izotopów argonu mierzono jednocześnie w okresie sześciu minut. Zmierzone zawartości izotopów skorygowano o puste linie ekstrakcyjne, które oznaczano przed każdą analizą próbki.
Interkalibrację detektora (IC) między różnymi kubkami Faradaya monitorowano (w Qtegra) co trzy dni przez przeskakiwanie pików 40 Ar. Współczynnik IC między H1 a CDD mierzono z nieznanymi za pomocą analizy online pipet powietrznych (wartości IC można znaleźć w Tabeli uzupełniającej 1 i https://doi.org/10.20383/103.0901 ). Wartość 295,5 została użyta dla stosunku atmosferycznego 40 Ar/ 36 Ar 42 na potrzeby rutynowego pomiaru dyskryminacji spektrometru masowego przy użyciu alikwotów powietrza i korekty dla argonu atmosferycznego w obliczeniach wieku 40 Ar/ 39 Ar. Korekty są dokonywane dla 40 Ar indukowanego neutronami z potasu, 39 Ar i 36 Ar z wapnia oraz 36 Ar z chloru 43 , 44 , 45 . Zbieranie danych i redukcję przeprowadzono przy użyciu Pychronu. Użyto stałych rozpadu zalecanych przez Min i innych 46 .
Markery molekularne
Około 30 g sproszkowanej próbki ekstrahowano w aparacie Soxhleta za pomocą DCM:MeOH (93:7) przez 72 godziny. Wyekstrahowany materiał organiczny (EOM) odparowano rotacyjnie do minimalnej objętości i przeniesiono do cylindra miarowego. Znany alikwot pobrano i odparowano do sucha pod stałym strumieniem azotu, a następnie zważono. Do pozostałej EOM dodano wzorce wewnętrzne (cholestan-d4, adamantan-d16, skwalan, naftalen-d8, fenantren-d10 i 1,1-binaftyl) na podstawie obliczonej suchej masy próbki. Węglowodory alifatyczne i aromatyczne frakcjonowano na kolumnie krzemionkowej i eluowano odpowiednio pentanem i alkoholem izopropylowym. W celu kontroli jakości w protokole uwzględniono ślepą próbę rozpuszczalnika, a jedną próbkę poddano duplikacji.
Frakcje węglowodorów alifatycznych i aromatycznych analizowano przy użyciu chromatografu gazowego (GC) Agilent 7890B sprzężonego ze spektrometrem masowym (MS) Agilent 5977 A. Analizę GC przeprowadzono przy użyciu kolumny kapilarnej HP-5MS (30 m × 0,25 mm × 0,25 μm), 1 µl próbki wstrzykiwanej w trybie splitless, temperatura wtryskiwacza 250 °C, gaz nośny hel przy stałej szybkości przepływu 1,0 ml/min. Temperaturę pieca GC utrzymywano początkowo na poziomie 40 °C przez 5 minut i ogrzewano z szybkością 4 °C/min do 325 °C, a następnie utrzymywano przez 15 minut. Spektrometr masowy pracował w trybie jonizacji elektronowej (EI) z energią elektronów 70 eV, pracując w trybie pełnego skanowania (50–550 mz) i wybranym monitorowaniu jonów (SIM).
Analizy izotopów węgla i tlenu stabilnego
Próbki z krawędzi policzkowej pobrano za pomocą diamentowej końcówki Dremel. Przetwarzanie odbyło się w Applied Geochemistry Group – Isotope Science Laboratory, Department of Earth, Energy and Environment, University of Calgary, Canada. Czyste gatunki minerałów węglanowych analizowano za pomocą ciągłej spektrometrii masowej izotopów przepływowych przy użyciu Thermo Finnigan GasBench® sprzężonego z DeltaVPlus®. Zważoną próbkę równą ~0,300 mg czystego kalcytu odważono do 12 ml Labco Exetainer (nr części: 038 W). Exetainery załadowano do poziomego stojaka bez zatyczki. Następnie ostrożnie wstrzyknięto około 200 µl specjalnie przygotowanego bezwodnego kwasu fosforowego tuż wewnątrz szyjki Exetainera. Ze względu na poziomą orientację kwas pozostaje w pobliżu szyjki fiolki w małej kropli, oddzielonej od próbki na dnie fiolki. Fiolki są trzymane poziomo, zamykane i przepłukiwane helem UHP przez 10 minut przy szybkości przepływu ~70 ml/min. Następnie fiolki są obracane pionowo, umożliwiając kroplom kwasu spływanie na dno fiolki i reagowanie z próbką. Następnie fiolki są umieszczane w podgrzewanym bloku GasBench w temperaturze 25 °C i pozostawiane do reakcji przez określony czas (minimum 5 h dla kalcytu). Następnie wydzielona przestrzeń nad cieczą CO2 jest automatycznie próbkowana przez ławkę gazową przy użyciu pętli próbki 50 µL i wlotu do źródła jonów spektrometru masowego w celu analizy stosunków 13 C/ 12 C i 18 O/ 16 O. Przestrzeń nad cieczą każdej fiolki jest próbkowana sześć razy przez wstrzyknięcie pętli. Pierwszy pik jest odrzucany, a 06_gb-delta_carbs kolejnych 5 wstrzyknięć jest pozyskiwanych. Jeżeli pierwszy pik jest >30 V, następne pięć wstrzyknięć jest automatycznie rozcieńczanych przez oprogramowanie w stosunku ~3:1.
Wybrane wewnętrzne materiały referencyjne laboratoryjne zostały uruchomione na początku i na końcu każdego zestawu próbek i są używane do normalizacji danych, a także do korygowania wszelkich dryftów przyrządów. Te wewnętrzne standardy laboratoryjne są okresowo kalibrowane względem międzynarodowych materiałów referencyjnych w celu zapewnienia dokładności skali Vienna Peedee Belemnite (VPDB). Wszystkie wyniki są podawane w notacji promilowej w odniesieniu do międzynarodowych skal VPDB odpowiednio dla δ 13 C i δ 18 O. Stabilne stosunki izotopów są wyrażane jako delta ( δ ) i są miarami różnicy „promila” (‰) lub części na tysiąc między stosunkiem izotopów próbki a znanym (międzynarodowym) materiałem standardowym. Wartości są podawane względem materiału referencyjnego VPDB dla węgla i tlenu. Stosunki izotopów tlenu są powszechnie porównywane zarówno do odniesień VSMOW 47 , jak i VPDB 48 . Tradycyjnie tlen w wodzie jest podawany w odniesieniu do VSMOW, podczas gdy tlen uwolniony ze skał węglanowych lub innych archiwów geologicznych jest podawany w odniesieniu do VPDB. Podobnie jak w przypadku wodoru, skala izotopów tlenu jest definiowana przez dwa materiały, VSMOW2 i SLAP2. Pomiary próbki δ 18 O względem VSMOW można przekonwertować na układ odniesienia VPDB za pomocą następującego równania: δ 18 O(VPDB) = 0,97001*δ 18 OVSMOW–29,99‰ .
Geochemiczna charakterystyka narzędzi kamiennych
Artefakty kwarcytowe z warstwy III ( n = 17) i warstwy IV ( n = 15) w wykopie 1 wybrano do pozyskania surowca zgodnie z rozważaniami stratygraficznymi, technologicznymi i litologicznymi. Przed analizą artefakty poddano indywidualnej sonikacji (10 minut; 40 kHz) w szklanych zlewkach przy użyciu wody destylowanej, aby ułatwić klasyfikację makroskopową i zapobiec zanieczyszczeniu geochemicznemu. Artefakty porównano z kolekcją odniesienia geologicznego i sklasyfikowano do dziewięciu grup makroskopowych, w tym Green1 ( n = 1), Gray1 ( n = 12), Gray2 ( n = 3), Red2 ( n = 1), Red6 ( n = 1), White1 ( n = 10), White2 ( n = 1), White3 ( n = 1) i White4 ( n = 2).
Artefakty charakteryzowano geochemicznie w sposób nieniszczący, kierując promieniowanie rentgenowskie na najbardziej płaskie powierzchnie, aby zmniejszyć rozpraszanie promieni rentgenowskich, przy użyciu spektrometru fluorescencji rentgenowskiej z dyspersją energii ARL QUANT’X firmy Thermo Scientific (okienko hamowania, lampa rentgenowska Rh o mocy 50 W, okienko berylowe o średnicy 76 μm, napięcie 4–50 kV, pompa próżniowa Edwards RV8). Natężenia promieniowania rentgenowskiego zostały automatycznie przeliczone na szacunkowe stężenia (główne tlenki = % wag.; pierwiastki śladowe = ppm) przy użyciu linii kalibracji najmniejszych kwadratów przeliczonej na rozproszenie Comptona na podstawie analizy trzynastu wzorców odniesienia, w tym AGV-2 (andezyt), BCR-2 (bazalt), BHVO-2 (hawait), BIR-1a (bazalt), GSP-2 (granodioryt), JR-1 (obsydian), JR-2 (obsydian), QLO-1 (latytu kwarcowego), RGM-2 (ryolit), SDC-1 (łupek łyszczykowy), STM-2 (syenit), TLM-1 (tonalit) i W-2a (diabaz). Docelowe pierwiastki obejmowały osiem głównych tlenków (MgO, Al 2 O 3 , SiO 2 , K 2 O, CaO, TiO 2 , MnO, Fe 2 O 3 ) i dziesięć pierwiastków śladowych (Cu, Zn, Rb, Sr, Y, Zr, Nb, Ba, Pb, Th). Każdy przebieg analityczny obejmował standard odniesienia RGM-2 w celu zapewnienia kalibracji urządzenia. Wszystkie stężenia głównych tlenków i większości pierwiastków śladowych (oprócz Cu i Ba) zarejestrowane dla standardu odniesienia RGM-2 mieściły się w dopuszczalnym zakresie w stosunku do zalecanych wartości.
Te same metody czyszczenia i warunki analityczne zastosowano w przypadku próbek geologicznych kwarcytu ( n = 333) pochodzących z siedmiu źródeł (Endonyo Okule, n = 18; Endonyo Osunyai, n = 27; Isilale Aratum, n = 6; Koonge, n = 21; Naibor Soit, n = 181; Naisiusiu, n = 51; i Oittii, n = 29), które stanowią zbiór referencyjny do celów pozyskiwania surowców. Próbki geologiczne podzielono na dwanaście grup makroskopowych, w tym Green1 ( n = 26), Gray1 ( n = 20), Gray2 ( n = 82), Red1 ( n = 12), Red2 ( n = 36), Red3 ( n = 38), Red4 ( n = 17), Red5 ( n = 3), White1 ( n = 38), White2 ( n = 12), White3 ( n = 35) i White4 ( n = 14).
Surowe szacunki stężenia dla artefaktów i próbek geologicznych znormalizowano przy użyciu metody skalowania Min-Max i posortowano do czterech makroskopowych grup (zielona, szara, czerwona, biała) w celu porównań geochemicznych. Czytelnik powinien zapoznać się z tekstem uzupełniającym, aby uzyskać dodatkowe informacje na temat standardu odniesienia RGM-2, zbioru odniesień geologicznych i identyfikacji źródła. Analizy statystyczne przeprowadzono przy użyciu algorytmu uczenia maszynowego nadzorowanego losowego lasu w RStudio przy użyciu siedmiu pakietów.
Metody palinologiczne
Trzydzieści jeden próbek przetworzono z piaszczystej gliny, mułowca i mułowca z Facji 2, 4 i 5, odpowiednio, do analizy palinologicznej zgodnie ze standardowymi technikami ekstrakcji w Geological Survey of Canada (Calgary). Proces obejmował mycie, trawienie kwasem, utlenianie roztworem Schulze’a i barwienie Safraniną O; pozostałości zamontowano za pomocą poliwinylu i płynnego bioplastiku. Obserwacje palynomorfów lądowych preparatów niesitowych przeprowadzono za pomocą mikroskopu światła przechodzącego Olympus BX61® z immersją olejową przy powiększeniu ×400 i ×1000. Obrazy cyfrowe wykonano za pomocą kamery Olympus DP72 i oprogramowania Stream Motion®. Osiemnaście próbek nie dało pyłku ani zarodników. Spośród produktywnych próbek suma pyłku i zarodników wahała się od 2 do 1623. Wszystkie próbki zawierały mikroskopijny węgiel drzewny. Obfitość względna pyłku i zarodników w produktywnych próbkach została obliczona przy użyciu sumy pyłków obligatoryjnie lądowych pyłków i zarodników, a obfitość mikroskopijnych węgli drzewnych i zarodników grzybów została obliczona na podstawie tej sumy. Stratygraficzny diagram pyłkowy został wykreślony przy użyciu programu komputerowego Tilia (Grimm, 1993–2001).
Analiza fitolitów
Ekstrakcję fitolitów zakończono w Clean Room Laboratory Uniwersytetu Calgary (Tropical Archaeology Laboratory, Earth Sciences Building, 811). Szczegółowe protokoły są dostępne w Federated Research Data Repository ( https://doi.org/10.20383/101.0123 ). Podsumowując, poszczególne próbki osadu przesiano przez sito o średnicy oczek 125 μm, z którego pobrano 3,0 g porcji osadu. Następnie przeprowadzono dyspersję gliny: osad zmieszano z 10 ml 0,1% heksametafosforanu sodu (NaPO3 ) 6 , stosując 15-minutową sonikację i nocne wytrząsanie orbitalne z prędkością 200 obr./min. Próbki przepłukano uprzednio zagotowaną zdejonizowaną wodą odwróconej osmozy (RODI) i suszono przez 2 dni w temperaturze >70 °C, a następnie zważono. Usuwanie substancji nieorganicznych polegało na tym, że próbki otrzymywały 10 ml równych części 3 N kwasu solnego (HCl) i kwasu azotowego (HNO 3 ), a następnie płukano, wirowano przy 3000 obr./min, suszono przez 2 dni w temperaturze >70 °C i ważono. Usuwanie substancji organicznych zostało zakończone przez dodanie 10 ml 30% nadtlenku wodoru (H 2 O 2 ) na gorącej płycie (70 °C), z płukaniem, suszeniem i ważeniem. Oddzielenie fitolitów nastąpiło przez dodanie 5 ml poliwolframianu sodu (3Na 2 WO 4 • 9WO 3 • H 2 O) o ciężarze właściwym 2,4. Próbki zawirowano i wirowano przy 3000 obr./min przez 5 min. Otrzymany supernatant przeniesiono do nowych probówek wirówkowych, a wodę użyto do stopniowego zmniejszenia ciężaru właściwego w dwóch dodatkowych frakcjach, koncentrując fitolity w trzeciej frakcji. Próbki przepłukano uprzednio zagotowanym RODI przed dwoma rundami wirowania przy 4500 obr./min i ostatecznym suszeniem (>70 °C) i ważeniem. Z tej ostatniej frakcji, alikwot 0,001 g przeniesiono za pomocą szklanej pipety na szkiełko mikroskopowe, zalano wodą i zbadano na świeżo, aby umożliwić obrót fitolity w celu klasyfikacji 3D. Mikroskopię wykonano przy ×40 przy użyciu Olympus BX51 (Olympus SC50, CellSens 2.0). Fitolity z każdej próbki liczono na trzech oddzielnych szkiełkach, każde o powierzchni 600 mm2 . Nomenklatura fitolity była zgodna z Madellą i innymi49 , z wyjątkami. Łączna liczba próbek osadów przetworzonych na potrzeby badania wyniosła 33, a łączna liczba odzyskanych fitolitów wyniosła 112.
Analiza fauniczna
Wszystkie okazy faunistyczne >2 cm analizowano za pomocą dobrze ugruntowanych metod faunistycznych. Metody te obejmują analizy całego zestawu próbek, oszacowania wieku poszczególnych osobników i modyfikacje tafonomiczne opisane poniżej.
Zarejestrowaliśmy element, zidentyfikowaną klasyfikację taksonomiczną, stronę (lewą lub prawą), część, twarz i wiek. Okazy pogrupowano w klasy wielkości, w tym bardzo duże (>1000 kg); duże (1000–300 kg), średnie (300–100 kg), a także małe (<100 kg). Częstotliwości względne, takie jak liczba zidentyfikowanych okazów (NISP), minimalna liczba elementów (MNE) i minimalna liczba osobników (MNI), zostały wykorzystane do oceny i porównania względnej liczebności elementów szkieletowych i gatunków 50 , 51 . Utworzyliśmy profile szkieletowe dla EN przy użyciu Minimalnych Jednostek Zwierzęcych (%MAU), obliczonych przy użyciu długich części kości MNE reprezentowanych w zespole. Wiek zwierząt oszacowano na podstawie wzoru wymiany i zużycia zębów 52 , przy czym okazy pogrupowano jako młode osobniki podrosłe (młode i podrosłe osobniki podrosłe), dorosłe (wczesne i późne dorosłe osobniki dorosłe), stare dorosłe i nieznane, których wieku nie można było oszacować. Podzieliliśmy klasy wiekowe na trzy główne klasy, aby utworzyć wykresy trójkątne53 i porównać je z innymi lokalnymi zbiorami danych.
Całkowitą powierzchnię wszystkich kości i zębów zbadano za pomocą stereomikroskopu (Opthec 120 Hz) 10−45x. Zespół wykazał różne modyfikacje tafonomiczne, które zostały ocenione i skategoryzowane na fazy depozycyjne odżywcze, subaerial i diageniczne. Ślady nacięć 54 w zespole są dominującą zaobserwowaną modyfikacją. Ślady nacięć w tym zespole obejmują ślady krojenia, ślady skrobania i ślady cięcia 55 . Mikroskopu Hirox KH 7800 użyto do obejrzenia okazów ze śladami nacięć, zmierzenia śladów nacięć i utworzenia profili śladów nacięć. Antropogeniczne złamania kości odnotowano w postaci dołków po uderzeniu, nacięć i otarć 56 . Zidentyfikowane ślady zębów obejmują dołki, nakłucia i bruzdy 57 na tkance gąbczastej. Zanotowano ich lokalizację na kości, a ich pomiary wykonano za pomocą suwmiarki cyfrowej 58 . W każdym z okazów stwierdzono obecność lub brak uszkodzeń subaeryjnych i diagenetycznych. Zarejestrowaliśmy wietrzenie 59 , deptanie 60 , 61 , ścieranie 62 , 63 , zaokrąglenie 64 , korozję chemiczną i rozpuszczanie.
Analiza litowa
Wszystkie narzędzia kamienne zostały zważone i zmierzone zgodnie z ich osią technologiczną. Informacje o surowcu i atrybuty techno-typologiczne zostały zarejestrowane w bazie danych. Surowce, rozróżniane makroskopowo, obejmowały kwarcyt, bazalt, fonolit, granitoid i piaskowiec. Próbki zostały sklasyfikowane do 10 grup technologicznych: manuporty , materiały udarowe, rdzenie, fragmenty rdzeni, odłupki, proksymalne odłamki, dystalne odłupki, retuszowane odłupki, LCT i nieokreślone odłamki. Rdzenie zostały sklasyfikowane według metody kucia, faset (liczby eksploatowanych powierzchni) i polarności (kierunku eksploatacji z platformy uderzeniowej).
Obserwowane przez nas metody redukcji rdzeni obejmują: (i) jednostronne jednobiegunowe podłużne: rdzenie z jedną powierzchnią eksploatowane z jednego kierunku; (ii) jednostronne dwubiegunowe przeciwstawne: rdzenie z jedną powierzchnią eksploatowane z dwóch przeciwnych kierunków; (iii) jednostronne dośrodkowe: rdzenie z jedną powierzchnią eksploatowane z co najmniej trzech kierunków zbiegających się w środku; (iv) dwustronne na kowadle: chociaż jest to technika kucia, a nie metoda, odnosi się do rdzeni z dowodami umieszczenia na pasywnej powierzchni; (v) dwustronne ortogonalne: rdzenie z dwiema przeciwległymi powierzchniami kutymi każdą z jednego kierunku; (vi) dwustronne dośrodkowe: rdzenie z dwiema powierzchniami eksploatacyjnymi zorganizowanymi w płaszczyznę poziomą, każda strona eksploatowana z co najmniej trzech kierunków zbiegających się w środku; (vii) wielostronne wielobiegunowe: rdzenie z czterema lub większą liczbą powierzchni eksploatowanych w czterech różnych kierunkach.
Retuszowana analiza płatków jest zgodna z analityczną typologią Laplace’a 65 . Gromadzenie danych odbywało się za pomocą programu Excel. Statystyki opisowe i wielowymiarowe prowadzono za pomocą oprogramowania do statystyki paleontologicznej. Dane użyte w PCA znormalizowano za pomocą wzoru Z-scores. Do generowania rysunków i wykresów użyto oprogramowania wektorowego Inkscape Open-Source.
Podsumowanie raportu
Więcej informacji na temat projektu badawczego można znaleźć w podsumowaniu sprawozdawczym Nature Portfolio Reporting Summary dostępnym pod tym artykułem.
Dostępność danych
Wszystkie dane niezbędne do wyciągnięcia wniosków z badania są zawarte w artykule. Zestawy danych geochronometrycznych i geochemicznych użyte do datowania i korelacji stratygraficznej, a także protokoły paleobotaniczne są publicznie dostępne w Canadian Federated Research Data Repository, a cyfrowe identyfikatory obiektów są cytowane w stosownych miejscach w artykule.
Odniesienia
-
Clark, D. Późnoplejstoceńskie kultury Afryki. Science 150 , 833–847 (1965).
-
McBrearty, S. & Brooks, AS Rewolucja, której nie było: nowa interpretacja pochodzenia współczesnego zachowania człowieka. J. Hum. Evol. 39 , 453–563 (2000).
-
Roberts, P. i Stewart, BA Definicja niszy „specjalisty ogólnego przeznaczenia” dla plejstoceńskiego Homo sapiens. Nat. Hum. Behav . 2 , 542–550 (2018).
-
Timmermann, A. i in. Wpływ klimatu na archaiczne siedliska ludzkie i sukcesje gatunków. Nature 604 , 495–501 (2022).
-
Zeller, E. i in. Adaptacja człowieka do różnych biomów w ciągu ostatnich 3 milionów lat. Science 380 , 604–608 (2023).
-
Foister, TIF, Žliobaitė, I., Wilson, OE, Fortelius, M. i Tallavaara, M. Homo heterogenus: zmienność w środowiskach Homo z wczesnego plejstocenu. Evol. Anth. 32 , 373–385 (2023).
-
Mercader, J. i in. Najwcześniejsze homininy Olduvai eksploatowały niestabilne środowiska ~ 2 miliony lat temu. Nat. Commun . 12 , 3 (2021).
-
Mussi, M. i in. Wczesny Homo erectus żył na dużych wysokościach i wytwarzał narzędzia zarówno oldowajskie, jak i acheulskie. Science 382 , 713–718 (2023).
-
Kleindienst, MR Wykopaliska na stanowisku JK2, wąwóz Olduvai, Tanzania. Quaternaria 17 , 145–208 (1973).
-
Leakey, MD, Olduvai Gorge: 5, wykopaliska w warstwach III, IV i warstwach Masek, 1968–1971 (Cambridge University Press, Cambridge, 1995).
-
Hay, RL Geologia wąwozu Olduvai: badanie sedymentacji w basenie półpustynnym (University of California Press, Berkeley, 1976).
-
Day, MH i Molleson, TI Zagadka z kości udowej JK2-A i fragmentu kości piszczelowej (OH 34) z wąwozu Olduvai w Tanzanii. J. Hum. Evol. 5 , 455–465 (1976).
-
Rightmire, GP Człowiekowate środkowoplejstoceńskie z wąwozu Olduvai w północnej Tanzanii. Amer . J Phys. Anth . 53 , 225–241 (1980).
-
Hay, RL Reakcje krzemianowe w trzech litofacjach półpustynnego basenu, wąwóz Olduvai, Tanzania. Min. Soc. Am. Spec. Pap. 3 , 237–255 (1970).
-
Stanistreet, IG i in. Zmieniające się środowiska depocentrów jeziora Palaeolake Olduvai i węglany jako horyzonty markerowe dla przerw i ekstremów na poziomie jeziora. Palaeogeogr. Palaeoclimatol. Palaeoecol . 560 , 110032 (2020).
-
Stanistreet, G. i in. Nowa stratygrafia basenu Olduvai i koncepcje stratygraficzne ujawnione przez rdzenie OGCP w depocentrum Palaeolake Olduvai w Tanzanii. Palaeogeogr. Palaeoclimatol. Palaeoecol . 554 , 109751 (2020).
-
Stanistreet, IG i in. Paleosalinność i paleoklimatyczne wskaźniki geochemiczne (pierwiastki Ti, Mg, Al) zmieniają się wraz z cyklicznością Milankovitcha (1,3 do 2,0 Ma), rdzenie OGCP, Palaeolake Olduvai, Tanzania. Palaeogeol. Palaeoclimatol. Palaeoecol . 546 , 109656 (2020).
-
Walker, TR Formowanie się czerwonych złóż na współczesnych i dawnych pustyniach. Geo. Soc. Am. Bull. 78 , 353–368 (1967).
-
Van Houten, FB Pochodzenie czerwonych zagonów – przegląd–1961–1972. Annu. Rev. Earth Planet. Sci. 1 , 39–61 (1973).
-
Deino, AL i in. Chronostratygrafia i modelowanie wieku rdzeni wiertniczych z plejstocenu z basenu Olduvai w Tanzanii (projekt rdzeniowania wąwozu Olduvai). Palaeogeogr. Palaeoclimatol. Palaeoecol. 571 , 109990 (2021).
-
Domínguez-Rodrigo, M. i in. Pierwszy częściowy szkielet Paranthropus boisei liczący 1,34 miliona lat z Bed II w wąwozie Olduvai w Tanzanii. PLoS Jeden . 8 , e80347 (2013).
-
McHenry, LJ, Njau, JK, de la Torre, I. i Pante, MC Geochemiczne „odciski palców” tufów Olduvai Gorge Bed II i implikacje dla przejścia oldowańskiego w acheulskie. Quat. Res. 85 , 147–158 (2016).
-
Thomas, DSG, red. Geomorfologia strefy suchej: proces, forma i zmiany na terenach suchych (Wiley-Blackwell, 2011).
-
White, F. Roślinność Afryki, opisowe wspomnienie towarzyszące mapie roślinności Afryki UNESCO/AETFAT/UNSO (UNESCO, 1983).
-
Huntley, BJ Ed. Środowisko Kuiseb: opracowanie bazowego punktu monitoringu. Raport komitetu ekosystemów lądowych. Raport nr 106 Południowoafrykańskie Narodowe Programy Naukowe (Narodowe Programy Naukowe: CSIR, 1985).
-
Bolinder, K. i in. Morfologia pyłku Ephedra (Gnetales) i jej implikacje ewolucyjne. Grana 55 , 24–51 (2016).
-
Barbolini, N. i in. „Ewolucja kenozoiczna biomu stepowo-pustynnego w Azji Środkowej”, Sci. Adv . 6 , eabb8227 (2020).
-
Freitag, H. i Maier-Stolte, M. Rodzaj przęśli w tropikalnej Afryce północno-wschodniej. Kew Bull. 58 , 415–426 (2003).
-
Caveney, S., Charlet, DA, Freitag, H., Maier-Stolte, M. i Starratt, AN Nowe obserwacje dotyczące chemii wtórnej świata Ephedra (Ephedraceae). Jestem. J.Bota. 88 , 1199–1208 (2001).
-
Ritchie, JC i Haynes, CV Strefowanie roślinności holoceńskiej w Saharze Wschodniej. Natura 330 , 645–647 (1987).
-
Uno, KT i in. Dieta ssaków i paleoekologia w okresie przejścia oldowańskiego w acheulskie w wąwozie Olduvai w Tanzanii na podstawie analizy stabilnych izotopów i zużycia zębów. J. Hum. Evol. 120 , 76–91 (2018).
-
Mees, F., Stoops, G., Van Ranst, E., Paepe, R. i Van Overloop, E. Charakter występowania zeolitów w złożach basenu Olduvai w północnej Tanzanii. Glina Górnik. 53 , 659–673 (2005).
-
Eckmeier, E. & Wiesenberg, GLB Krótkołańcuchowe n- alkany (C16-20) w starożytnej glebie są użytecznymi markerami molekularnymi spalania biomasy w okresie prehistorycznym. J. Arch. Sci. 36 , 1590–1596 (2009).
-
Nicholas, CJ i in. Stratygrafia i sedymentologia górnej kredy do paleogenu Kilwa Group, południowe wybrzeże Tanzanii. J. Afr. Earth Sci. 45 , 431–466 (2006).
-
Gentry, W. & Gentry, A. Fossil Bovidae (Mammalia) of Olduvai Gorge, Tanzania. Część II, (Londyn, 1978).
-
Pante, MC Większy zespół skamieniałości ssaków z JK2, Bed III, Olduvai Gorge, Tanzania: implikacje dla zachowań żywieniowych Homo erectus. J. Hum. Evol. 64 , 68–82 (2013).
-
Yravedra, J. Rubio-Jara, S., Courtenay, LA i Martos, JA Rzeź ssaków przez Homo erectus na stanowisku acheulskim z dolnego plejstocenu w Juma’s Korongo 2 (JK2), warstwa III, wąwóz Olduvai, Tanzania. Quat. Sci. Rev . 249 , 106612 (2020).
-
Miall, AD Aktualizacja uniformitarianizmu: Stratygrafia jako zbiór „zamrożonych wypadków”. Geo. Soc. Spec. Publ. 404 , 11–36 (2015).
-
Dazé, A., Lee, JKW i Villeneuve, M. Badanie interkalibracyjne standardów sanidynu i biotytu 40Ar/39Ar z Fish Canyon oraz kilka uwag na temat wieku tufu z Fish Canyon. Chem. Geol. 199 , 111–127 (2003).
-
Kuiper, KF i in. Synchronizacja zegarów skalnych historii Ziemi. Science 320 , 500–504 (2008).
-
Phillips, D., Onstott, TC & Harris, JW 40 Ar/ 39 Ar datowanie metodą sondy laserowej inkluzji diamentowych z Premier kimberlitu. Nature 340 , 460–462 (1989).
-
Steiger, RH i Jager, E. Podkomisja ds. geochronologii: konwencja dotycząca stosowania stałych rozpadu w geo-i kosmochronologii opracowana przez. Earth Planet. Sci. Lett. 36 , 359–362 (1977).
-
Roddick, JC Wysoka precyzja interkalibracji standardów 40Ar-39Ar. Geochim. Cosmochim. Acta 47 , 887–898 (1983).
-
Renne, PR i in. Interkalibracja standardów, wieków bezwzględnych i niepewności w datowaniu 40Ar/39Ar. Chem. Geol. 145 , 117–152 (1998).
-
Renne, PR i Norman, EB Oznaczanie okresu półtrwania 37Ar metodą spektrometrii masowej. Phys. Rev. C. 63 , 473021–473023 (2001).
-
Min, K., Mundil, R., Renne, PR i Ludwig, KR Test na błędy systematyczne w geochronologii 40 Ar/ 39 Ar poprzez porównanie z analizą U/Pb ryolitu 1,1-Ga. Geochim. Cosmochim. Acta 64 , 73–98 (2000).
-
Hut, G. Spotkanie grupy konsultantów ds. próbek referencyjnych izotopów trwałych do badań geochemicznych i hydrologicznych. (Międzynarodowa Agencja Energii Atomowej, Wiedeń, 1987).
-
Dansgaard, W. Stabilne izotopy w opadach atmosferycznych. Tellus 16 , 436–468 (1964).
-
Madella, M., Alexandre, A. i Ball, T. Międzynarodowy kod nomenklatury fitolitów 1.0. Ann. Bot. 96 , 253–260 (2005).
-
Binford, LR Bones: starożytni ludzie i mity współczesne. (Academic Press, 1981).
-
Gray, JS Wzory liczebności gatunków. Symposia Br. Ecol. Soc. 27 , 53–68 (1987).
-
Bunn, HT i Pickering, TR Zalecenia metodologiczne dotyczące analiz śmiertelności kopytnych w paleoantropologii. Quat. Res. 74 , 388–394 (2010).
-
Steele, TE i Weaver, TD Modyfikowany wykres trójkątny: udoskonalona metoda porównywania profili śmiertelności w próbkach archeologicznych. J. Archaeol. Sci. 29 , 317–322 (2002).
-
Potts, R. i Shipman, P. Ślady nacięć wykonanych narzędziami kamiennymi na kościach z wąwozu Olduvai w Tanzanii. Nature 291 , 90–108 (1981).
-
Lyman, RL Jednostki ilościowe i terminologia w zooarcheologii. Am. Antiquity 59 , 36–71 (1994).
-
Blumenschine, RJ i Selvaggio, MM Ślady perkusji na powierzchniach kości jako nowa diagnostyka zachowań hominidów. Nature 333 , 763–765 (1988).
-
Binford, LR Rzeźnictwo, dzielenie się i zapis archeologiczny. J. Anthropol. Archaeol. 3 , 235–257 (1984).
-
Andrés, M., Gidna, AO, Yravedra, J. i Domínguez-Rodrigo, M. Badanie różnic wymiarowych śladów zębów (wgłębień i nacięć) na kościach zmodyfikowanych przez małe i duże drapieżniki. Archaeol. Anthropol. Sci. 4 , 209–219 (2012).
-
Behrensmeyer, AK Informacje tafonomiczne i ekologiczne z wietrzenia kości. Paleobiologia 4 , 150–162 (1978).
-
Fiorillo, AR Tafonomia kamieniołomu Hazard Homestead (grupa Ogallala), hrabstwo Hitchcock, Nebraska. Współprac. Geol. 26 , 57–97 (1988).
-
Reynard, JP Deptanie w miejscach przybrzeżnych: badanie eksperymentalne nad wpływem muszli na kości w osadach przybrzeżnych. Quat. Int. 330 , 156–170 (2014).
-
Mózg, CK Wietrzenie kości i problem pseudonarzędzi kostnych. S. Afr. J. Sci. 63 , 3 (1967).
-
Shipman, P. & Rose, J. Wczesne zachowania hominidów związane z polowaniem, rzeźnictwem i przetwarzaniem tusz: podejścia do zapisu kopalnego. J. Anthropol. Archaeol. 2 , 57–98 (1983).
-
Olsen, SL i Shipman, P. Modyfikacja powierzchni kości: deptanie kontra rzeźnictwo. J. Archaeol. Sci. 15 , 535–553 (1988).
-
Laplace, G. La typologie analytique etstructuree: base rationnelle d’étude des industriallithiques et osseuses. Colloques nationaux du CNRS Banques de données archéologiques, 92–143 (1972).
Podziękowanie
Po pierwsze, autorzy doceniają istotny wkład w wiedzę przedstawioną w niniejszym dokumencie społeczności Masajów w wąwozie Oldupai. Ponadto wielu studentów studiów podyplomowych i współpracowników pomagało w logistyce terenowej, uzyskiwaniu pozwoleń i wykopaliskach archeologicznych. Tanzania Commission for Science and Technology oraz Ministry of Natural Resources and Tourism zezwoliły na prace na tym terenie, a licencje na eksport próbek pochodziły z wydziału ds. zabytków, a tanzański sekretarz wykonawczy z Mining Commission. Nasza praca była sponsorowana przez Canadian Social Sciences and Humanities Research Council w ramach jej Partnership Grant Program kierowanego przez pierwszego autora/PI (JM). Profesor Noah McLean pomagał w modelowaniu głębokości wieku. Profesor Maxine Kleindienst pomagała w kontekstualizacji swoich wykopalisk z lat 60.
Deklaracje etyczne
Konflikty interesów
Autorzy deklarują brak konfliktu interesów.
Recenzja ekspercka
Informacje o recenzji eksperckiej
Communications Earth & Environment dziękuje Gabriele Macho i innym anonimowym recenzentom za ich wkład w recenzję ekspercką tej pracy. Główni redaktorzy: Jiaoyang Ruan i Carolina Ortiz Guerrero. [Dostępny jest plik recenzji eksperckiej].
Informacje dodatkowe
Uwaga wydawcy: Springer Nature zachowuje neutralność w kwestii roszczeń jurysdykcyjnych na opublikowanych mapach i powiązań instytucjonalnych.